Самые легкие цветы из бумаги. Своими руками пошагово + 232 фото
cамые простые цветы из бумаги
Хотите научиться делать цветы из бумаги своими руками и ищете простые варианты для начинающих? Тогда здесь вы найдете то, что вам необходимо. Посмотрите примеры, как сделать легкие цветы из бумаги своими руками – простые, недорогие и их можно использовать для разных событий и мероприятий.
Для того, чтобы сделать простой цветок, подойдет практически любая бумага – гофрированная, креповая, салфетки, даже картон и кофейные фильтры. Если вы знакомы с техникой оригами, то здесь найдете три примера, как сделать легкий цветок оригами. Есть много методов и инструментов, чтобы сделать простые цветы из бумаги, все зависит только от того, насколько вы креативны. Вот 15 проектов простых бумажных цветов, которые вы можете попробовать сделать своими руками и научить ваших детей.
Содержание:
- Как сделать легкую розу из бумаги своими руками
- Легкие объемные цветы из бумаги
- Простые цветы из салфеток своими руками
- Легкие цветы из цветной бумаги
- Как сделать из бумаги самый легкий цветок
- Цветы с конфетой из салфеток
- Простой цветок оригами своими руками
- Легкий тюльпан оригами
Как сделать легкую розу из бумаги своими руками
Эта инструкция, как сделать легкую розу из цветной бумаги. Задача не сложная и предназначена для начинающих, просто вам нужно использовать представленные на сайте бесплатные шаблоны. Необходимое время: 25 минут. Вы можете выбрать любую цветную бумагу, которую хотите. На каждый цветок вам понадобится 4 розовых 5-лепестковых слоя и 1 зеленый чашелистик. Бесплатный шаблон находится в файлах PDF и SVG. Используйте край ножниц, чтобы изогнуть лепесток, попробуйте свернуть обе стороны. Затем приклейте провод к первому слою. Немного клея на кончике проволоки поможет закрепить стебель. Свернув форму пятиугольника внизу каждого слоя, вы получите точную форму цветка. После этого вырежьте шаблон чашелистика из зеленой бумаги и приклейте в нижней части цветка.
Бесплатный шаблон находится в файлах PDF и SVG. Используйте край ножниц, чтобы изогнуть лепесток, попробуйте свернуть обе стороны. Затем приклейте провод к первому слою. Немного клея на кончике проволоки поможет закрепить стебель. Свернув форму пятиугольника внизу каждого слоя, вы получите точную форму цветка. После этого вырежьте шаблон чашелистика из зеленой бумаги и приклейте в нижней части цветка.
Оберните небольшую полоску бумаги, чтобы сформировать емкость. Теперь сам цветок. Здесь можно сделать 2 типа бутона розы. Для первого потребуется только 1 слой (5 лепестков). А второму бутону розы понадобится 2 слоя (равняется 10 лепесткам). Разрежьте лист по шаблону. Использовать технику в видео на сайте, чтобы обрезать край листа, что поможет сделать его более реалистичным.
Необходимые материалы:
- Цветная бумага
- Ножницы или автомат для резки
- Клеевой пистолет или клейкий клей
- Маленький стилус
Автор: Анна.
Источник фото: www.dreamyposy.com/how-to-make-paper-rose/
Простые объемные цветы из бумаги
Пошаговое руководство, как сделать простые бумажные цветы. Сначала вырежьте из картона круг. Размер должен быть примерно таким, каким вы хотите, чтобы был ваш цветок (размер не так важен, так как вы всегда можете приклеить другой бумажный кружок, если он маленький). Нарисуйте на бумаге несколько квадратов, которые немного длиннее с одной стороны и вырежьте их (это должны быть внутренние лепестки, и поэтому они самые маленькие). Теперь обрежьте у квадратов один конец. Если предпочитаете, можете нарисовать их или сделать шаблон на листе бумаги. Используйте карандаш, чтобы округлить лепестки, как показано на рисунке. Затем начинайте приклеивать лепестки к кругу.
Когда используете все лепестки, сделайте еще, на этот раз немного больше. Помните: если у вас заканчивается свободное пространство на нижнем круге, вы можете просто вырезать больший круг и указать его в нижней части старого круга – все равно никто не увидит на готовом цветке.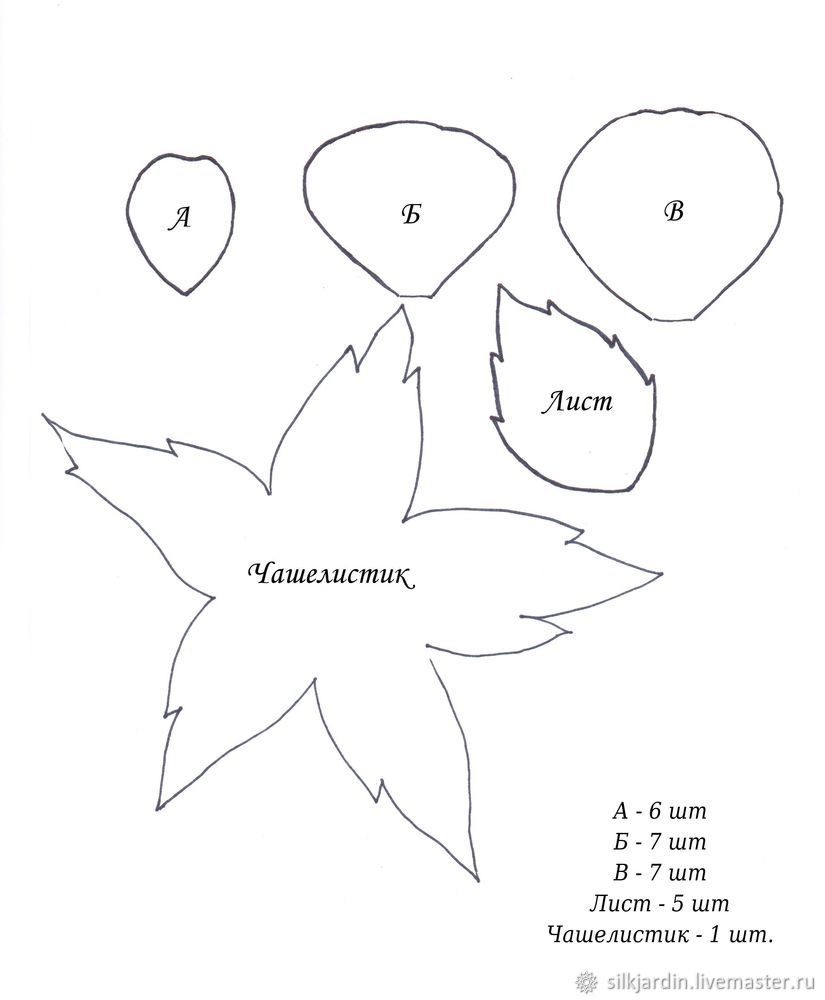 Подождите, пока клей высохнет, и наслаждайтесь готовым цветком. Если хотите, чтобы цветок был более прочным, покройте его лаком для волос. Чем больше слоев лака, тем жестче становится бумага. Таким образом, вы сможете использовать цветок, например, для брошки.
Подождите, пока клей высохнет, и наслаждайтесь готовым цветком. Если хотите, чтобы цветок был более прочным, покройте его лаком для волос. Чем больше слоев лака, тем жестче становится бумага. Таким образом, вы сможете использовать цветок, например, для брошки.
Источник фото: www.instructables.com/id/Paper-Craft-Flowers/
Цветок из салфеток по шаблону
Этот вариант тоже не сложный, но немного хлопотней. Чтобы сделать цветок из салфеток своими руками, сначала распечатайте шаблон, ссылка на сайт подл галереей. Вы можете изменить размеры лепестков, манипулируя изображением в вашей любимой программе или изменив размер на принтере (если принтер имеет функцию масштабирования). Вы можете вырезать лепестки все сразу или по отдельности. После этого сложите их вместе, так чтобы центры совпадали. Хотя они не должны полностью совпадать. Сделайте отверстие и пропустите провод через все лепестки.
Для того, чтобы сделать стебель, возьмите цветочный провод и большой шарик (пластмассовый шарик или бусину), согните провод пополам и закрепите шарик на одном конце. Протолкните к центру, и крутить проволоку, как показано на фото. Затем нанизайте лепестки на стебель. При использовании нескольких размеров, убедитесь, что самые маленькие лепестки нанизаны на первом слое, а затем идут более крупные. Используйте немного клея, чтобы держать каждый лепесток на месте. Вы можете согнуть и сложить лепестки, чтобы придать больший объем и форму цветка.
Протолкните к центру, и крутить проволоку, как показано на фото. Затем нанизайте лепестки на стебель. При использовании нескольких размеров, убедитесь, что самые маленькие лепестки нанизаны на первом слое, а затем идут более крупные. Используйте немного клея, чтобы держать каждый лепесток на месте. Вы можете согнуть и сложить лепестки, чтобы придать больший объем и форму цветка.
После того, как клей высохнет, используйте цветочные ленты, чтобы покрыть проволочный стебель. Намотайте плотно вокруг всего ствола, чтобы покрыть весь провод. Ваш цветок из салфеток готов и можно собрать в букет или поставить в вазу, или можете скрутить проволоку вокруг запястья, получится красивый браслет! Для работы вам понадобятся: цветные салфетки, проволока, шаблоны лепестков, цветочная лента, острогубцы (для скручивания проволоки, но это по желанию, это легко можно сделать пальцами, острогубцы нужны, если делаете большое количество цветов).
Источник фото: craftinomicon.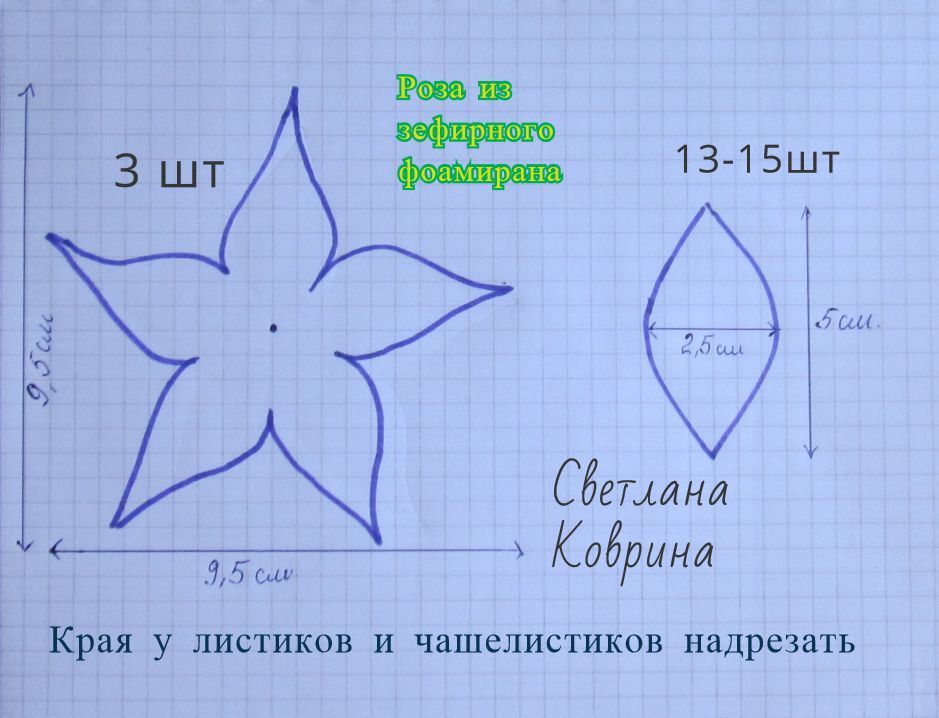 blogspot.com/2012/01/more-tissue-paper-flowers.html
blogspot.com/2012/01/more-tissue-paper-flowers.html
Легкие цветы из цветной бумаги
Эти красивые цветы из бумаги действительно легко сделать. Вам понадобится всего 2 квадратных кусочка желтой бумаги, 1 квадратный лист зеленой бумаги, клей для рукоделия и ножницы. Сложите желтую бумагу дважды и вырежьте 4 квадратных кусочка. Затем сложите один из маленьких квадратов в треугольник 3 раза. Повторите столько раз, сколько хотите сделать лепестков. Отрежьте нижнюю часть для центрального отверстия. Для первого выреза нужна всего одна прорезь по центру. Склейте вырезы вместе. Потом склейте все вместе от большого к маленькому, с самым маленьким, находящимся в центре. Дайте цветку высохнуть и приклейте кусочек зеленой бумаги к стеблю. С помощью клея соедините цветок со стеблем. Соберите букет, используя разные цвета.
Источник фото: www.instructables.com/id/Paper-Flowers-5/
Самый легкий цветок из бумаги
Это самый простой цветок из бумаги, который можно сделать за несколько минут. Все, что вам нужно это, папиросная бумага, или салфетки (подойдет и гофрированная бумага), ножницы, цветочная проволока. Для этих маленьких цветков использовались 4 листа тонкой бумаги. Сложите их поверх друг друга и соберите гармошкой. Найдите середину. Используя кусок цветочного провода приблизительно 15 – 18 см в длину (также сложенные пополам) закрепите бумагу, то есть просто оберните проволокой в центре. Затем, все, что вам нужно сделать, это мягко потянуть за каждый слой бумаги по мере необходимости и распушить. Вот и все, самый легкий цветок из бумаги готов.
Все, что вам нужно это, папиросная бумага, или салфетки (подойдет и гофрированная бумага), ножницы, цветочная проволока. Для этих маленьких цветков использовались 4 листа тонкой бумаги. Сложите их поверх друг друга и соберите гармошкой. Найдите середину. Используя кусок цветочного провода приблизительно 15 – 18 см в длину (также сложенные пополам) закрепите бумагу, то есть просто оберните проволокой в центре. Затем, все, что вам нужно сделать, это мягко потянуть за каждый слой бумаги по мере необходимости и распушить. Вот и все, самый легкий цветок из бумаги готов.
Источник фото: craftinomicon.blogspot.com/2012/01/tissue-paper-flowers.html
Цветы с конфетой из салфеток
Эти красочные цветы из салфеток легкие в изготовлении, красивые и включают конфеты — так что это идеальный вариант на день Святого Валентина. Чтобы сделать цветок, вам понадобится:
- Цветные салфетки
- Ножницы
- Дырокол
- Леденцы на палочке
- Степлер
- Бамбуковые шампуры
- Зеленая или цветочная лента
Как сделать цветок из салфеток с конфетой
- Разрежьте девять 20-ти см салфеток
- Положите квадраты друг на друга, чередуя цвета.

- Согните один край в 3-х сантиметровую полосу.
- Переверните стопку и откиньте край назад в противоположную сторону. Продолжайте складывать гармошкой.
- Обрежьте оба конца в изогнутую форму.
- Растяните гармошку и найдите складку, которая ближе всего к середине. Повторно сложите вдоль этой складки и пробейте отверстие по краю, чтобы у вас получилось отверстие рядом с центром квадрата.
- Вставьте леденец в перфорированное отверстие, затем замените складки гармошкой.
- Скрепите с обеих сторон палочку леденца.
- Осторожно разделите слои салфеток по обеим сторонам леденца на палочке, распушив по мере создания формы цветка.
- Если положите леденцы на палочке в вазу, используйте зеленую ленту, чтобы прикрепить палочку на бамбуковом шампуре и продолжайте оборачивать ленту, чтобы создать стебель цветка.

Источник фото: www.goodhousekeeping.com/holidays/valentines-day-ideas/a42463/diy-tissue-paper-flower-lollipops/
Как сделать простой цветок оригами
Здесь вы узнаете, как сложить простой цветок оригами, который красиво смотрится в букетах из бумажных цветов! Сложите бумагу пополам по диагонали, в обе стороны.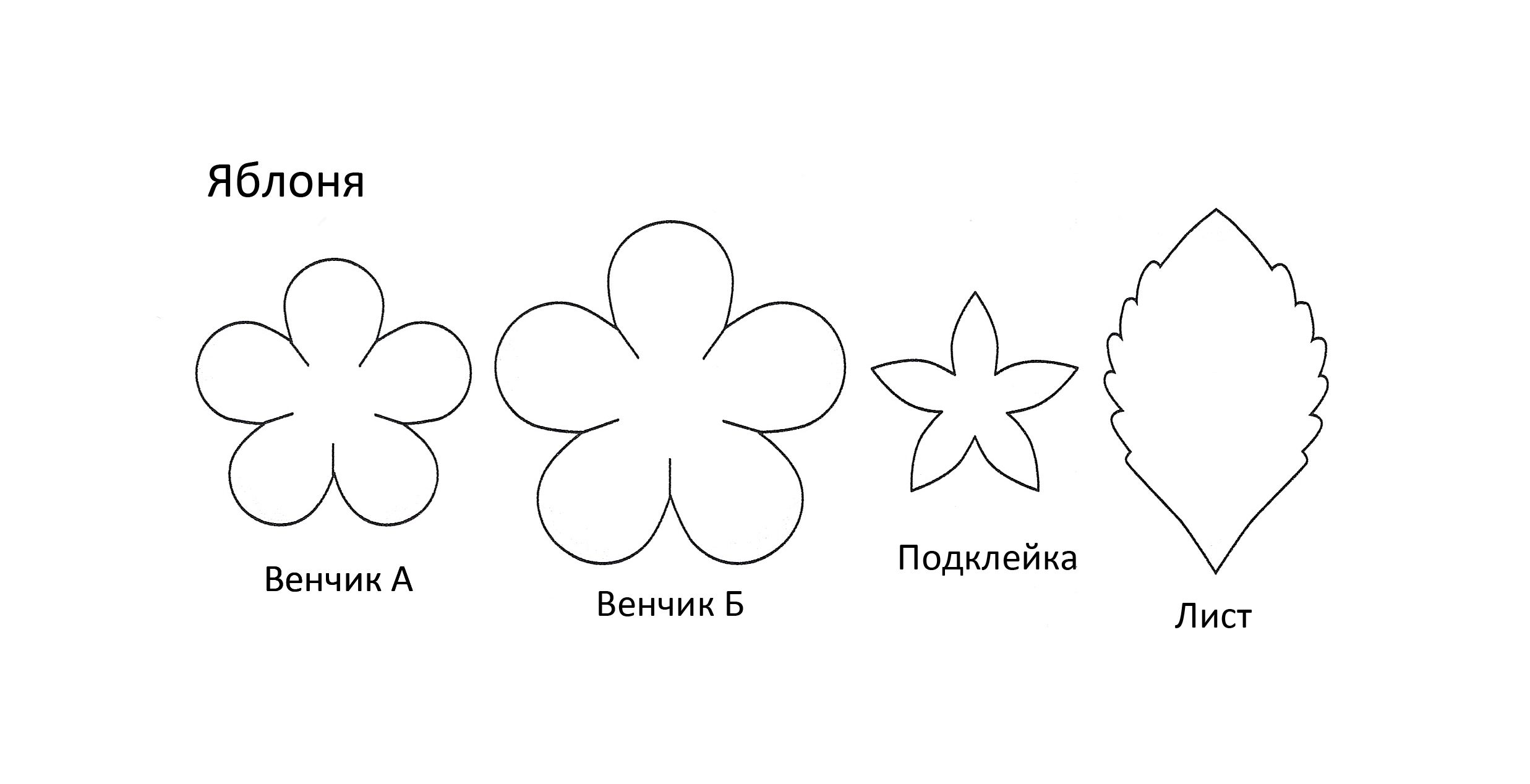 Сложите все углы к середине. Повторите. Затем сложите пополам в обе стороны, чтобы придать форму. Назовем сторону, которую только что свернули, «верхней стороной», а сторону с двумя наборами углов, сложенными к середине, «нижней стороной». Верхняя сторона должна быть обращена вверх, а нижняя — вниз.
Сложите все углы к середине. Повторите. Затем сложите пополам в обе стороны, чтобы придать форму. Назовем сторону, которую только что свернули, «верхней стороной», а сторону с двумя наборами углов, сложенными к середине, «нижней стороной». Верхняя сторона должна быть обращена вверх, а нижняя — вниз.
Посмотрите на закрылки на нижней стороне. Вставьте в них пальцы и вытолкните все к середине. Сложите обе части на нижнюю сторону и согните. Переверните и откиньте два клапана, пока не откроются четыре маленьких в центре. Сложите назад и заправьте, чтобы в центре появился белый квадрат. Переверните и откиньте клапаны, на этом простой цветок оригами завершен.
Источник фото: www.wikihow.com/Fold-a-Simple-Origami-Flower
Простой цветок оригами своими руками
Цветок оригами твист — это цветок необычной формы, который складывается с использованием техники оригами. Это немного необычный дизайн, на фото вы видите витой стебель и красивый бумажный цветок.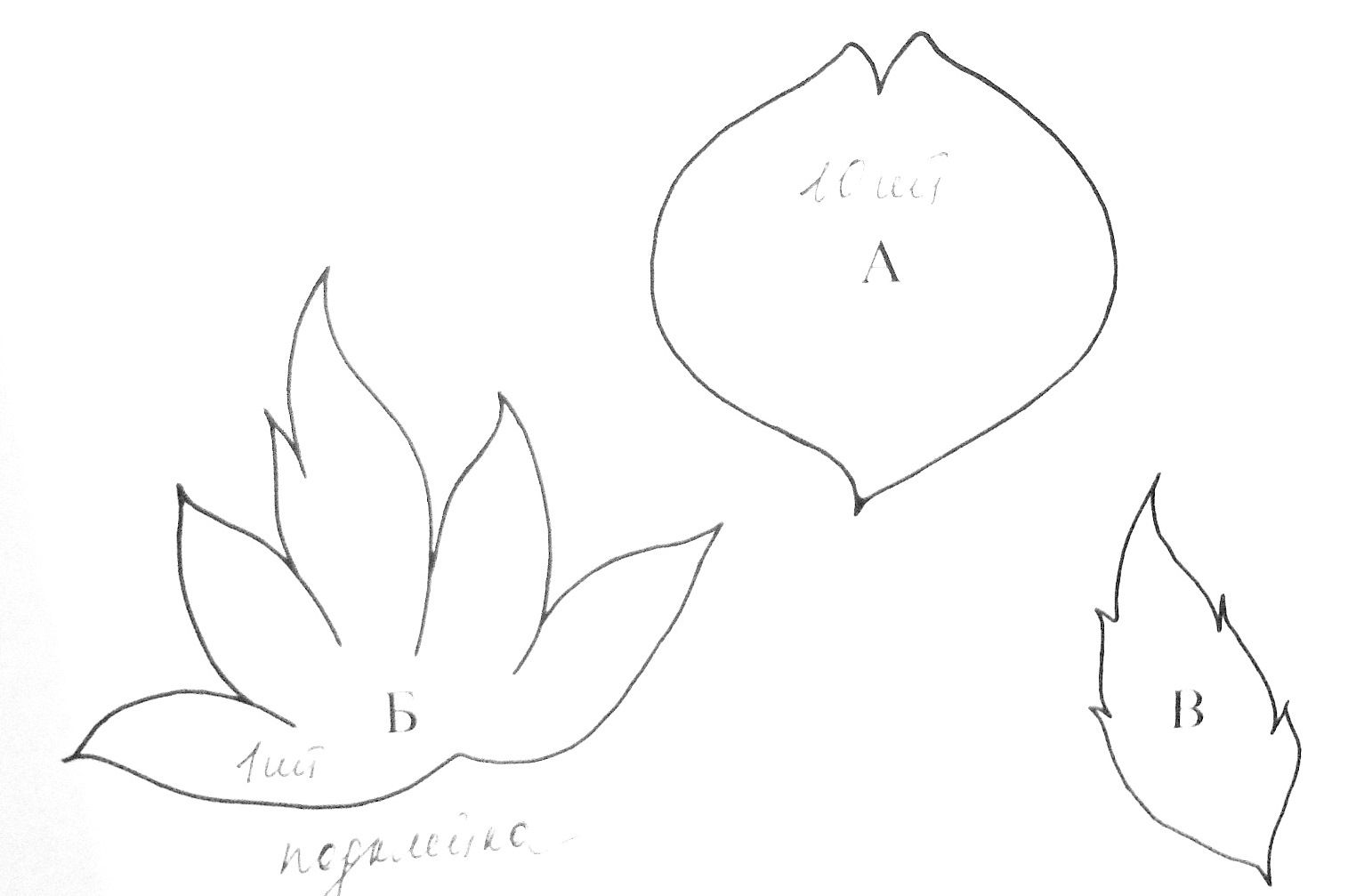 Эти цветы оригами можно использовать как украшение или для чего-нибудь другого, что вам больше нравится. Мастер класс представленный от сайта wikihow вы можете посмотреть ниже на пошаговых фото. Инструкцию можно прочитать на сайте, ссылка под галереей.
Эти цветы оригами можно использовать как украшение или для чего-нибудь другого, что вам больше нравится. Мастер класс представленный от сайта wikihow вы можете посмотреть ниже на пошаговых фото. Инструкцию можно прочитать на сайте, ссылка под галереей.
Источник фото: www.wikihow.com/Make-an-Origami-Twist-Flower
Как сделать легкий тюльпан оригами
Инструкция для тех, кто хочет научиться складывать легкий тюльпан оригами с двумя листьями.
Это простая поделка для начинающих, так что, пошаговых фото будет достаточно, чтобы сложить этот цветок оригами.
Источник фото: www.origami-flower.org/flower-origami-tulip-easy.php
Как сделать мини цветы из салфеток
Ищете идею оформления бюджетной вечеринки? Посмотрите, как сделать мини цветы из салфеток своими руками. Приготовьте 5 салфеток, сложите в стопку и вырежьте центр по длине. Это будет заготовка для двух цветков. Сложите салфетку гармошкой, со складками приблизительно 3 см. Обвяжите посередине нитками или шпагатом. Обрежьте концы по форме лепестка. Разделите слои салфеток и «распушите в центре». Идите по кругу, пока все 5 слоев не будут распушены. Салфетка очень легко рвется, поэтому будьте осторожны, но даже если немного порвете, не волнуйтесь. Никто никогда не заметит! Повторите со второй стопкой салфеток, чтобы сделать второй цветок! Вуаля! У вас есть 2 красивых миниатюрных цветка из салфеток!
Обвяжите посередине нитками или шпагатом. Обрежьте концы по форме лепестка. Разделите слои салфеток и «распушите в центре». Идите по кругу, пока все 5 слоев не будут распушены. Салфетка очень легко рвется, поэтому будьте осторожны, но даже если немного порвете, не волнуйтесь. Никто никогда не заметит! Повторите со второй стопкой салфеток, чтобы сделать второй цветок! Вуаля! У вас есть 2 красивых миниатюрных цветка из салфеток!
Источник фото: hellocreativefamily.com/how-to-make-diy-mini-tissue-paper-flowers-party-decorations/
Водяная лилия из гофрированной бумаги
Если ищете инструкции, как сделать водяные лилии из гофрированной бумаги, посмотрите этот пример. Техника не сложная: сформируйте лепесток и склейте его на круговую основу слоями. Это руководство будет очень простым. Расходные материалы: гофрированная бумага, ножницы, клей. Еще приготовьте небольшой кусочек картона для основания — нарезать на круг диаметром приблизительно 5 см.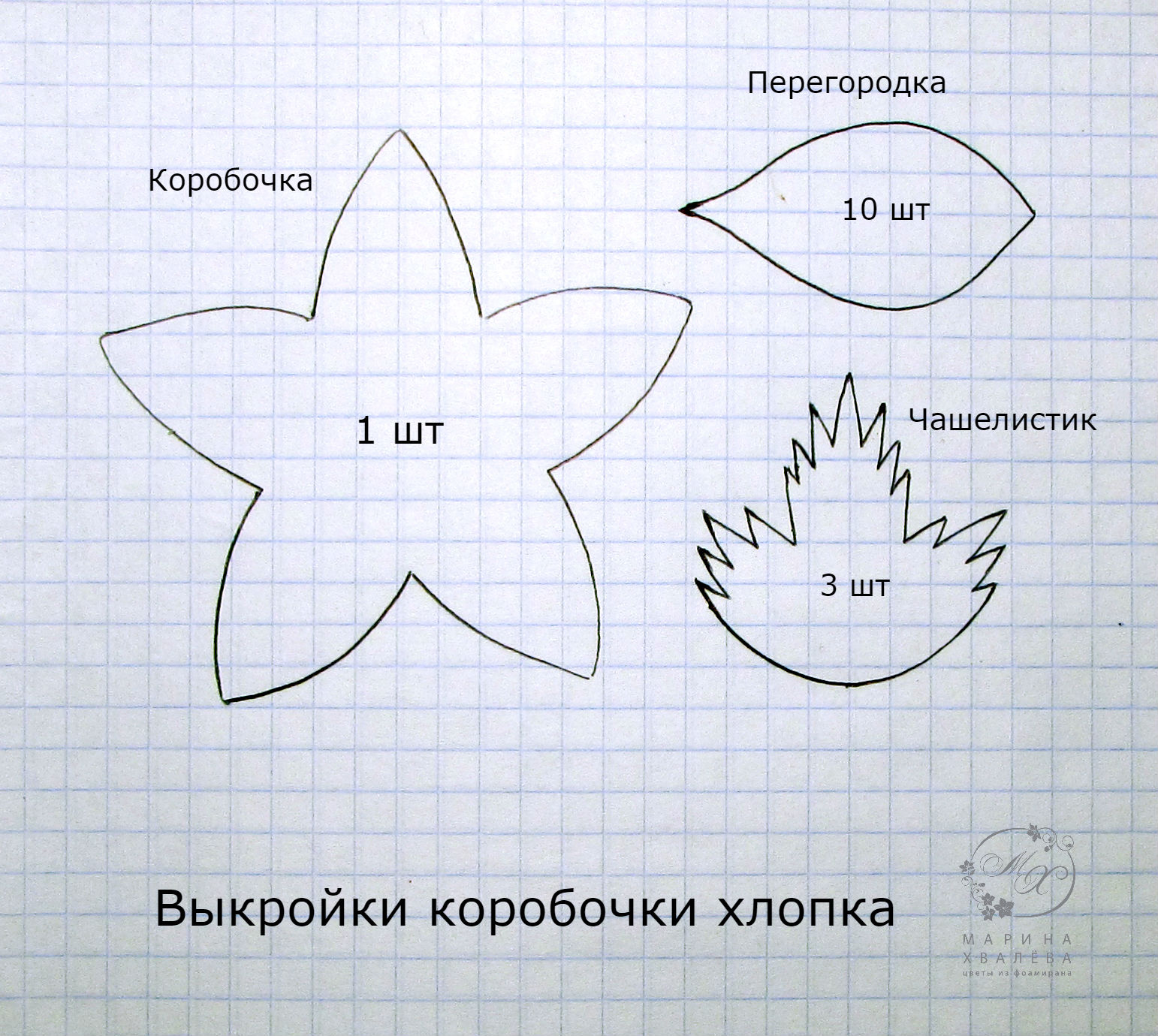
Сначала нужно нарезать гофрированную бумагу на полоски. Вам понадобится около 20 полос. Когда вы продвигаетесь слоями вверх, обрежьте лепестки так, чтобы самые широкие находились снаружи, а самые узкие внутри по центру. Используя подушечку большого пальца, вытяните гофрированную бумагу у основания лепестка, прежде чем она начнет сужаться. Лепесток должен иметь форму чаши, но не нужно делать этого для лепестков в первом слое. Сделайте небольшую прорезь у основания лепестка, где на фото нарисована черная пунктирная линия. Склейте один клапан над другим. Чем больше вы перекрываете закрылки, тем прямее будет лепесток. Наклейте наружный слой на основание круга. Добавьте второй слой лепестков. Можете добавить столько слоев, сколько захотите. Чтобы сделать центр, понадобится полоса бумаги, шириной примерно 8 см и высотой 1,5 см. Осторожно сделайте полосу с надрезами на 3/4.
Пальцами слегка сверните край от себя. Затем скатайте полоску бумаги вверх и нанесите немного клея. Раздвиньте края, чтобы создать центр, а затем приклейте его к цветку. И вот водяная лилия из гофрированной бумаги готова! БОНУС: Используя тот же метод, также можете сделать цветок лотоса из гофрированной бумаги. Просто нужно обрезать лепестки так, чтобы они были шире и короче лепестков водяной лилии. Если у вас есть какие-либо вопросы по пошаговым фото, полную инструкцию можно прочитать на сайте.
И вот водяная лилия из гофрированной бумаги готова! БОНУС: Используя тот же метод, также можете сделать цветок лотоса из гофрированной бумаги. Просто нужно обрезать лепестки так, чтобы они были шире и короче лепестков водяной лилии. Если у вас есть какие-либо вопросы по пошаговым фото, полную инструкцию можно прочитать на сайте.
Источник фото: ashandcrafts.com/crepe-paper-water-lily/
Цветы гиацинта из цветной бумаги
Хороший способ поднять настроение в холодные осенние дни, это сделать красивые цветы из цветной бумаги, например, гиацинты! Ничуть не хуже роз или тюльпанов. Они выглядят немного утомительно, но сделать гиацинты из бумаги не так уж и сложно. К тому времени, когда вы сделаете штуки три, то потом вам нужно будет всего около 5 минут на цветок. И главное, что выглядят эти бумажные цветы так ярко и весело, что это определенно стоит усилий! Мастер класс посмотрите ниже на пошаговых фото, если возникнут вопросы, подробная инструкция на сайте всегда в вашем распоряжении.
Автор: Дебби Чапман
Источник фото: onelittleproject.com/paper-hyacinth-flowers/?utm_source=kidsstuffworld.com&utm_medium=referral&utm_campaign=pubexchange_module
Василек с конфетой из гофрированной бумаги
Для цветка вам понадобятся: гофрированная бумага синего цвета, серебристая бумага с металлическим оттенком для конфеты. Васильки из бумаги можно сделать легко и быстро. Вырежьте 3 заготовки из синей гофрированной бумаги для лепестков (7 х 7 см). Вырежьте красивую каемку по верхнему краю с помощью ножниц для кутикулы. Потом сделайте разрезы, как показано на фото. Растяните лепестки посередине, придайте им форму лодки.
Сделайте небольшие порезы на каждом лепестке приблизительно 5 мм. Из проволоки изготовьте заготовку для конфеты. Чтобы обернуть конфету, нам понадобится серебристая мятая бумага с металлическим оттенком. Вырежьте заготовку 10 х 10 см и заверните конфету. Зафиксируйте первую линию лепестков ниткой. Затем, во втором ряду, вставьте лепестки между лепестками первого ряда. Ваш василек из бумаги с конфетой готов. Наслаждайтесь!
Затем, во втором ряду, вставьте лепестки между лепестками первого ряда. Ваш василек из бумаги с конфетой готов. Наслаждайтесь!
Источник фото: candy-bouquets-ideas.com/hand-made-cornflowers-a-tutorial
Цветок из фильтров от кофе
Цветы из кофейных фильтров сделать не сложнее, чем из обычной бумаги. Вот основные вещи, которые вам понадобятся: кофейные фильтры, ножницы с несколькими, скотч, тонкие деревянные палочки или провод. Цветок состоит из двух частей – помпона в центре и лепестков. Сделать цветок просто и недорого, посмотрите пошаговые фото, на сайте есть инструкция, так что в ваших руках и удачной поделки.
Источник фото: greenweddingshoes.com/diy-paper-flower/
Цветы магнолии из бумаги
Цветущие деревья, это лучшее, что дарит нам весна. Их цвет объявляет о том, что зима закончилась и голые ветки покрываются красивыми цветами. Даже после того, как они закончили цветение, опавшие лепестки украшают землю. К сожалению, цветущие ветки деревьев долго в вазе не простоят. Этот проект разработан, чтобы вы могли самостоятельно могли сделать простые цветы из бумаги, в данном случае цветущие ветви магнолии. Они будут цвести для вас всю весну или даже несколько лет, если захотите.
К сожалению, цветущие ветки деревьев долго в вазе не простоят. Этот проект разработан, чтобы вы могли самостоятельно могли сделать простые цветы из бумаги, в данном случае цветущие ветви магнолии. Они будут цвести для вас всю весну или даже несколько лет, если захотите.
Итак, сначала вырежьте два куска из белой и светло-розовой крепированной бумаги. Распылите с каждой стороны аэрозольный клей (делайте это на улице или в хорошо проветриваемом месте над газетой) Сожмите две части вместе, убедившись, что складки бумаги лежат в одном направлении. Сделайте несколько мазков на розовой стороне крепа. Это нужно для акцентирования основания лепестков. Дайте высохнуть.
Вырежьте лепестки из креповой бумаги с зерном, идущим вертикально. Каждому цветку нужно 6 лепестков. Аккуратно вытяните центр и обожмите основание каждого.
Согните конец провода более чем пару раз с помощью плоскогубцев. Оберните два лепестка вокруг провода, лицом друг к другу, с помощью цветочной ленты. Добавьте еще два лепестка лицом друг к другу, перекрывая первые два, а затем добавьте еще два. Аккуратно сложите лепестки наружу. Оберните провод вокруг ветки и обрежьте излишки. Для более мелких цветков, согните проволоку около основания цветка и обрежьте проволоку. Нанесите на ветку горячий клей. Отрежьте пучок листьев, около 1,5 см длиной. Закрепите горячим клеем листья вокруг оснований каждого цветка. Добавьте пару маленьких пучков листьев и лепестков на ветку, чтобы получились бутоны. Расставьте ветки в простой вазе и наслаждайтесь!
Добавьте еще два лепестка лицом друг к другу, перекрывая первые два, а затем добавьте еще два. Аккуратно сложите лепестки наружу. Оберните провод вокруг ветки и обрежьте излишки. Для более мелких цветков, согните проволоку около основания цветка и обрежьте проволоку. Нанесите на ветку горячий клей. Отрежьте пучок листьев, около 1,5 см длиной. Закрепите горячим клеем листья вокруг оснований каждого цветка. Добавьте пару маленьких пучков листьев и лепестков на ветку, чтобы получились бутоны. Расставьте ветки в простой вазе и наслаждайтесь!
Источник фото: www.vitaminihandmade.com/2014/05/diy-paper-magnolia-blossoms.html
Вот еще несколько фото очень легких и простых цветов из бумаги
Клубника из фоамирана
Автор:
Клубника из фоамирана
- В мастер-классе «Клубника из фоамирана своими руками» Ольга Сыротюк показывает новичкам, как сделать ягоды, цветочки и листики клубники для создания композиции.
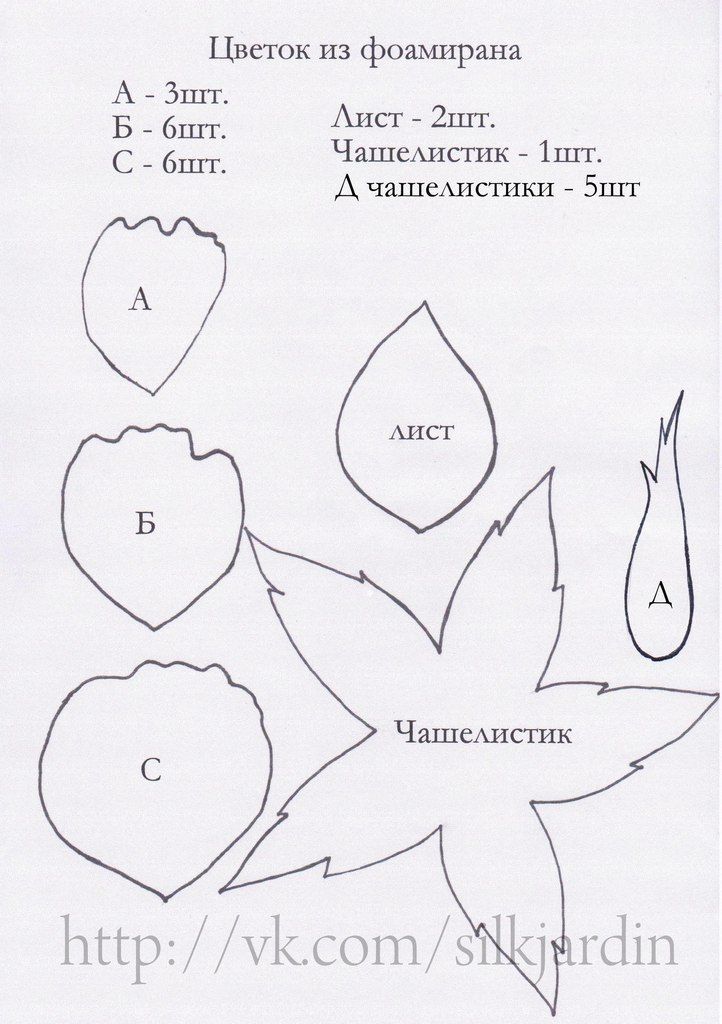
Для изготовления ягоды клубники из фоамирана нам понадобится:
Фоамиран красного, зеленого цвета (пр-во Иран, 0,8 мм)
Шаблон ягоды, цветка, чашелистика и листа клубники
Бисер
Игла с нитью
Канцелярский нож или ножницы
Специальный инструмент для изготовления цветов или изогнутая спица
Подушка для формирования листиков или поролоновая губка, обтянутая хлопчатобумажной тканью
Термоклей
Наполнитель для ягоды
Утюг
- Фото 1. На бумаге рисуем эскиз ягод, цветов и листочков клубники.
- Фото 2. Шаблон ягоды кладем на красный фоамиран и вырезаем нужное для композиции количество заготовок ягодок.
- Фото 3. Из зеленого фоамирана вырезаем необходимое количество заготовок листиком и чашелистиков.
- Фото 5. Заготовки ягод прикладываем к горячему утюгу и, как только заготовка слегка округлилась, сразу убираем.
- Фото 6. Прикладываем к утюгу цветы клубники, лепестки расправляем в нужном направлении.
 Прикладываем к утюгу цветы клубники, лепестки расправляем в нужном направлении.
Прикладываем к утюгу цветы клубники, лепестки расправляем в нужном направлении.- Фото 7. У заготовок чашелистиков делаем насечки.
- Фото 8. Прикладываем к утюгу.
- Фото 9. Складываем лепестки.
- Фото 10. И скручиваем верх листиков чашелистика.
- Фото 11. И расправляем заготовку.
- Фото 12-16. На заготовки клубнички пришиваем бисер.
- Фото 17. Обратная сторона заготовки.
- Фото 18. Закончили вышивку.
- Фото 19. Делаем для цветочка серединку из бисера: в центр заготовки вводим иглу ч ниткой.
- Фото 20. Нанизывем на нитку 5 бисеринок, вводим в центр иглу и протягиваем нитку на обратную сторону.
- Фото 21. Одна петелька из бисера готова.
- Фото 22-23. Аналогично делаем вторую и третью петельки и серединка готова.
Чтобы дочитать статью до конца, перейдите на следующую страницу ниже
2 мастер-класса с пошаговыми фото
В этом мастер-классе будем своими руками учится делать яркий летний цветок – подсолнух из фоамирана. Сделаем два вида подсолнуха полевой из иранского и садовый из зефирного фоамирана. Цветок будет довольно крупный на стебле, поэтому использовать его лучше для составления цветочных композиций. Подсолнухи хорошо сочетаются с полевыми цветами, часто их сочетают с ирисами, ромашками, фрезией и хризантемой из фоамирана. При желании вы сможете сделать подсолнух поменьше и закрепить его на заколке для волос.
Содержание:
Полевой подсолнух из иранского фоамирана: МК
Фото готового подсолнуха из фоамирана на стебле.
Материалы для изготовления подсолнуха
Как всегда начинаем наш МК с составления списка необходимых материалов для нашего солнечного цветка.
Иранский фоамиран:
- желтый, оливковый, коричневый или черный;
Материалы для тонировки:
- оранжевая, зеленая пастель, а также губка для нанесения пастели и растушевки;
Материалы для обработки и сборки подсолнуха:
- молд каттлеи;
- утюг;
- клей секундный;
- ножницы зигзаг и обычные;
- проволока №28-30 и 1 мм;
- тейп лента.
Шаблоны для подсолнуха
Подсолнух будем делать без отдельной выкройки. Для приготовления шаблонов для изготовления подсолнуха берем полоску желтого фоамирана шириной 5,2 и вырезаем прямоугольники шириной 2,2 см 20 штук в виде капелек.
И вторую полоску 6 см. из нее вырезаем прямоугольники шириной 2,5 см – 20 штук. Берем полоску оливкового фоамирана 60 см* 3,5 см., полоску черного 70* 4 см.
Тонировка лепестков
Все лепестки тонируем оранжевой масленой пастелью. Тонировку наносим с двух сторон.
Шаблоны для сердцевины подсолнуха
Черную полоску нарезаем заборчиком, не дорезая до края 0,8 см.
Полоску из оливкового фоамирана нарезаем бахромой, но неочень мелко.
Нарезанную бахрому нагреваем на утюге при температуре шелк-шерсть, так чтобы кончики чуть загнулись.
Так же обрабатываем и нарезанный заборчик, чтобы кончики загнулись как реснички.
Обработка лепестков и сборка полевого подсолнуха
Все лепестки обрабатываем одинаково. Нагреваем на утюге при температуре шерсть-шелк и обрабатываем на молде хорошо продавливая текстуру.
Затем берем проволоку делаем петельку и начинаем туго накручивать бахрому и подклеивать. Получился диаметр серединки 3,5 см.
Затем на зеленую бахрому начинаем накручивать черную полоску заборчика. Так чтобы заборчик закрывал зеленую серединку. Так же подклеиваем местами.
Потом берем маленькие лепестки и подклеиваем их на черную полоску.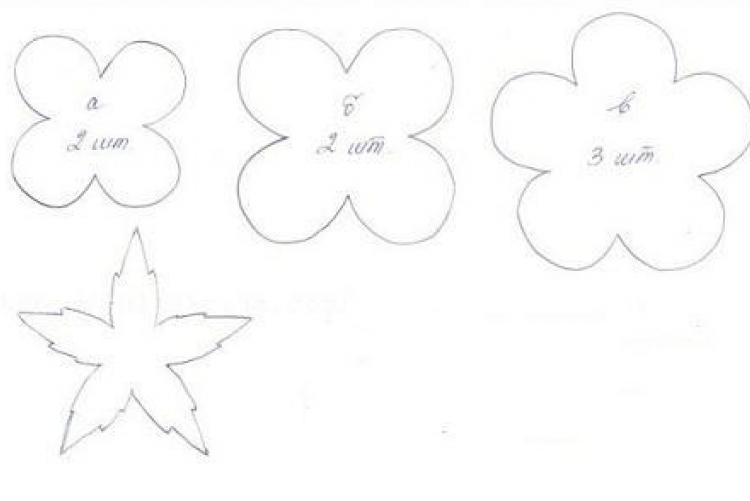 Так чтобы их было видно.
Так чтобы их было видно.
Подклеив первый ряд маленьких лепестков, начинаем приклеивать второй в шахматном порядке. То есть лепестки второго ряда клеим между лепестками первого.
Затем в шахматном порядке приклеиваем большие лепестки.
Делаем листья, чашелистик и стебель для подсолнуха
Из оливкового фоамирана вырезаем два круга диаметром 9 и 10 см. И из них вырезаем чашелистики в виде солнышка.
Наносим не большую тонировку с одной стороны масленой оливковой пастелью.
Затем обрабатываем на утюге.
Затем подклеиваем чашелистик к цветку подсолнуха, подклеиваем в начале лучики через один , затем остальные при необходимости формируем складочки. Утолщаем ножку подсолнуха. Сегодня утолщение ножки я сделала с помощью трубочек из газет. Скрутила трубочки промазала их клеем ПВА и вставила туда проволоку, на которую собирали подсолнух и даем немного подсохнуть.
Затем надрезаем второй чашелистик одеваем на стебель и тоже подклеиваем по кругу и в руке немного прижимаем.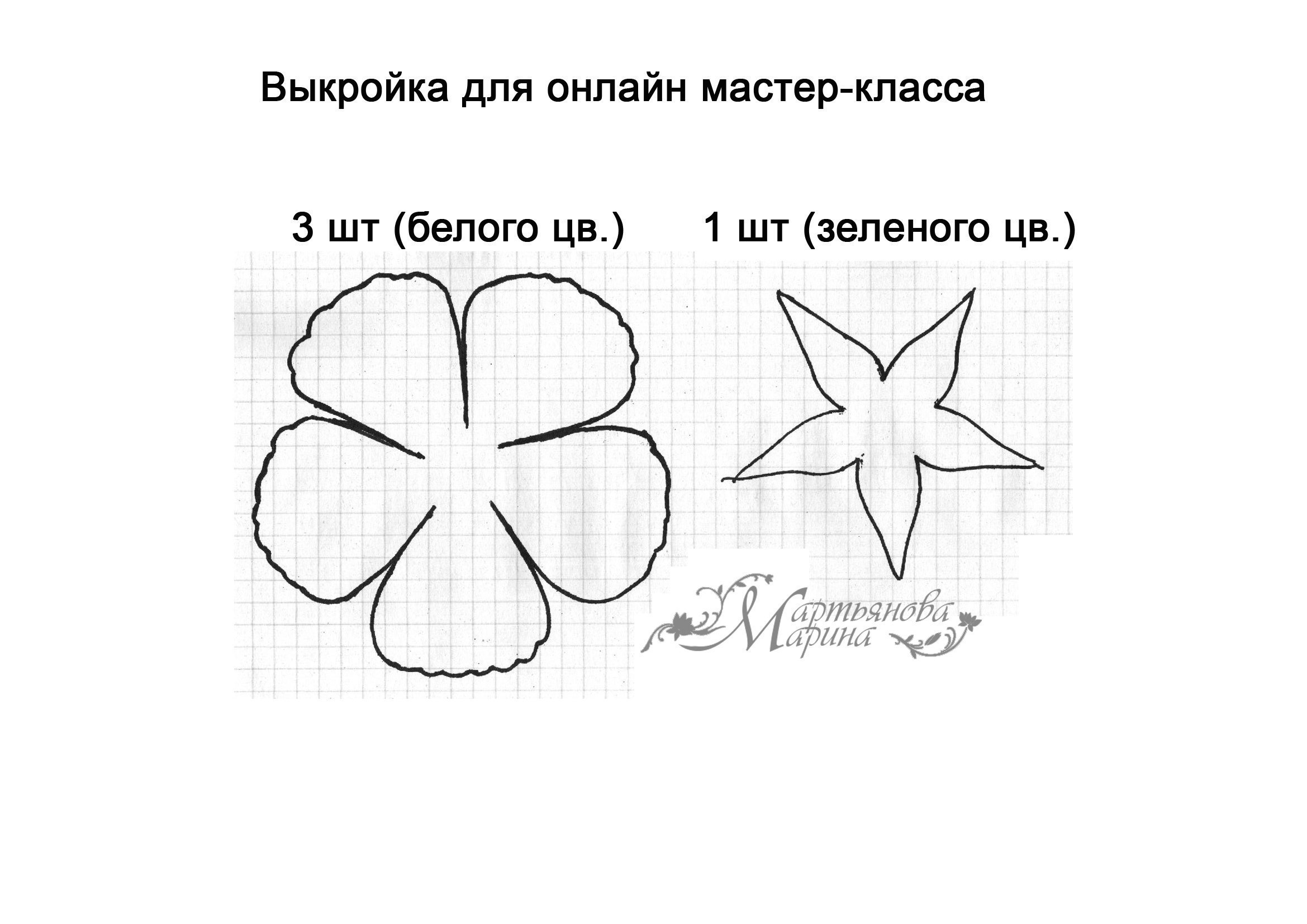
Из оливкового фоамирана вырезаем несколько листочков, можно разного размера. Палочкой наносим прожилки.
Для выделения текстуры нанесем тонировку оливковой постелью.
Для придачи листьям реалистичности обработаем листочки на утюге, на краях листочков сделаем не большие волны.
Приклеим листочки на литоны и обработаем литоны тейп лентой.
Обработаем стебель подсолнуха тейп лентой, при этом закрепим листочки на стебель. Нагибаем немного головку самого подсолнуха Вот и все.
Вот такой подсолнух из иранского фоамирана получился.
Садовый подсолнух из зефирного фоамирана: пошаговые фото
Этот вид подсолнуха делается несколько иначе, чем предыдущий. У него более тонкие, изогнутые лепестки и совсем другая сердцевинка.
Фото садового подсолнуха из зефирного фоамирана.
Для данного МК необходимы следующие материалы:
Зефирный фоамиран:
- желтый двух оттенков;
Иранский фоамиран:
- черный, оливковый;
Материалы для тонировки:
- оранжевая, зеленая пастель плюс губка для тонировки;
Материалы для обработки и сборки подсолнуха:
- утюг;
- моментальный клей;
- ножницы обычные;
- проволока №28-30 и 1 мм;
- тейп лента;
- флокс пудра;
- акриловый лак.
Готовим шаблоны для цветка
Подсолнух будем делать без отдельной выкройки. Для приготовления шаблонов для изготовления подсолнуха берем полоску желтого зефирного фоамирана шириной 6,5 см и вырезаем прямоугольники шириной 3,5 см 30-25 штук в виде капелек.
И вторую полоску 4,5 см. Из нее вырезаем прямоугольники шириной 2,5 см – 20-15 штук.
Берем полоску черного фоамирана 60 см* 7 см.
Разрезаем данную полоску пополам и каждую половинку складываем в 3 раза и проклеиваем.
Для проклеивания полосок я использовала клей “Кристалл”.
Затем полоски нарезаем на бахрому. 12 см первой полоски нарезаем бахрому шириной 1,2 мм., затем бахрому режим 2,5 см.
Вторую полоску мы нарезаем бахромой 2,5 мм полностью.
Для стебля подсолнуха берем обыкновенный провод, если есть потолще.
Так как у меня провод недостаточно толстый я делаю утолщение из бумажных салфеток. То есть обматываю и обклеиваю провод.
Берем полоску оливкового иранского фоамирана 6,5 см и нарезаем ее на прямоугольники 3,5 см. Затем из них вырезаем капельки 15-20 штук.
Обрабатываем лепестки для садового подсолнуха
Берем листок офисной бумаги и раскладываем на ней лепестки ,накрываем другим листом бумаги и хорошенько проглаживаем их утюгом с двух сторон. Чтобы наши листочки стали тоненькими.
Маленькие лепестки обрабатываем также.
Большие лепестки тонируем с одной стороны оранжевой масленой пастелью полностью.
Маленькие лепестки тонируем оранжевой масленой пастелью с двух сторон.
Затем лепестки хорошенько перетираем между пальчиками. Чуть расправляем середину лепестков кончики оставляем перекрученными.
Большие лепестки также перетираем между пальчиками и середину лепестка вытягиваем делая чашечку. Также обрабатываем лепестки чашелистика, но предварительно нагрев их на утюге при температуре шелк- шерсть, хорошенько перетирая между пальчиками. Сформировав лепесток, снова нагреваем кончик лепестка и еще раз его перекручиваем.
Собираем середину садового подсолнуха
Берем полоску зефирного желтого фоамирана, ширина полоски зависит от вашего дырокола. У меня дырокол 1,8 см. Необходимо надыроколить 35 шт таких цветочков.
Маленькие цветочки перетираем между пальчиками.
Берем подготовленный стебель и начинаем накручивать и подклеивать бахрому черную с 1,2 мм. Формируем середину на одном уровне. Так подклеиваем обе черные полоски. Серединку с обратной стороны для крепкости заливаем клеем из горячего пистолета.
Затем серединку немного тонируем желтой и зеленой пастелью.
Затем пинцетом вклеиваем перетертые маленькие цветочки. Вклеиваем там где бахрома немного отходить.
Пошаговая сборка цветка
Начинаем сборку подсолнуха с маленьких лепестков. Приклеиваем их на расстоянии 0,5 см. друг от друга. Низ лепестка немного прищипываем.
Второй ряд маленьких лепестков приклеиваем между лепестками первого ряда. Лепестки приклеиваем также с небольшими защипами.
Большие лепестки приклеиваем по кругу чуть-чуть внахлест.
Второй ряд больших лепестков приклеиваем между лепестками первого ряда больших лепестков, в шахматном порядке.
Берем полоску оливкового фоамирана шириной 0,7-0,5 см и перетираем ее между пальчиками, затем расправляем и обрабатываем ей стебель подсолнуха.
Берем зеленые лепестки и обклеиваем чашеложу подсолнуха, формируя чашелистик. Первый ряд лепестков приклеиваем по кругу в плотную к стеблю. Второй ряд лепестков приклеиваем в шахматном порядке.
Как сделать листья?
Из зеленого фоамирана иранского вырезаем листья. Палочкой наносим текстуру листа. Масленой зеленой пастелью наносим тонировку и хорошо растушевывая.
Берем литоны и приклеиваем к листочкам. Краешки листочков нагреваем на зажигалке и вытягиваем.
Берем акриловый лак и флокс пудру наносим на чашелистик . После подсыхания лишнее стряхиваем.
Обрабатываем стебель тейп лентой и одновременно крепим листочки. Затем слегка промазываем стебель и низ листочков акриловым и посыпаем флокс пудрой. После подсыхания все лишнее стряхиваем.
Вот такой чудесный садовый подсолнух садовый получился. его можно использовать в свадебных или интерьерных композициях.
Мастер-класс. Венок с цветами персика из фоамирана.
Сегодня мы хотим вам показать МК по цветам персика из корейского фоамирана. Вы можете использовать элементы цветов для изготовления заколки, броши, бутоньерки.
Материалы: фоамиран персиковый, шампань, тычинки мелкие «топленое молоко», кусочек глины, проволока 24, шаблон цветка, утюг, супер-клей, ножницы, стек с шариками, шаблон листа
Краска 11, 20, 23, 39, 40
1. На проволоку №24 при помощи кусочка глины фиксируем несколько тычинок.
2. Нанесите краску №23 на пальчик и легонько дотроньтесь до верхушки тычинок.
3. Вырежете по шаблону 2 цветка, сначала затонируйте цветок краской №20, затем высветлите серединку краской №40.
4. Приложите лепесток к утюгу.
5. Пока лепесток теплый, согните его гармошкой и прокрутите пальчиками. Обработайте так каждый лепесток.
6. Нагрейте стек на утюге и обработайте края лепестка.
7. Зафиксируйте получившийся цветок и тычинки на супер-клей.
8. Приклейте второй цветок с небольшим сдвигом.
9. Затонируйте фоамиран цвета шампань краской №39.
10. Вырежьте чашелистики по шаблону и затонируйте краской №23.
11. Приложите чашелистик к утюгу, для придания формы.
12. Уберите ножницами 2 сектора от большого чашелистика и приложите его к утюгу.
13. Зафиксируйте на клей ПВА маленький чашелистик.
14. Приклейте второй чашелистик сверху, фиксируя сектора между секторами первого чашелистика, имитирую почку.
15. Затонируйте стебель и чашелистик краской №23
16. Приложите листик к утюгу.
17. Пока листик теплый, согните его гармошкой и прокрутите пальчиками.
18. Приклейте сначала один листик, затем второй.
19. Приклейте чашелистик к листикам, имитируя почку.
20. Соберите венок при помощи тейп-ленты по своему усмотрению.
Наш венок готов!
Как сделать лаванду из фоамирана своими руками
Лаванда очень популярна благодаря своим декоративным качествам, ее невысокие кустики выглядят очень нежно и изящно. Из фоамирана тоже можно сделать веточки лаванды, которые будут смотреться довольно реалистично.
Лаванда из фоамирана – пошаговый мастер-класс (мк)
Для изготовления лаванды ним понадобится
- сиреневый фоамиран;
- розовый фоамиран;
- зеленый фоамиран;
- тейп-лента;
- проволока;
- клей универсальный прозрачный;
- ножницы.
Порядок работы
- Сделаем выкройку лаванды – вырежем из бумаги маленький цветок лаванды, деталь чашелистиков в виде звездочки и узкий вытянутый лист.
- Вырежем цветки лаванды из фоамирана. Из сиреневого фоамирана вырежем 16 цветочков, а из розового – 10 цветочков.
- Вырежем чашелистики и листья из зеленого фоамирана. Нам понадобится пять деталей чашелистиков и два листа.
- Цветки сомнем, для этого лепестки каждого цветка соберем вместе и покрутим в руках.
- В середину каждого цветка капнем клей и снова покрутим в руках, чтобы цветок превратился в бутончик. Один цветочек оставим не склеенным.
- Чашелистики сомнем, для этого каждую деталь чашелистика сложим так, чтобы лучики оказались вместе и покрутим в руках.
- Расправим чашелистики, немного вытягивая лучики в стороны.
- К каждой детали чашелистиков приклеим по пять цветочков.
- Возьмем проволоку длиной 20 см и обмотаем ее тейп-лентой. В центре одного цветка сделаем отверстие, наденем его на проволоку, смажем клеем его серединку и склеим лепестки вместе.
- В центре одной детали чашелистиков сделаем отверстие, наденем деталь на проволоку и приклеим.
- Таким же образом приклеим к проволочному стеблю остальные чашелистики с цветочками.
- Листья скрутим, прокручивая их в руках.
- Немного расправим листья, вытягивая их вверх.
- Приклеим листья к стеблю и примотаем их тейп-лентой.
Лаванда из фоамирана — шаблон
Веточка лаванды готова. Букетик из таких веточек будет хорошо смотреться в небольшой изящной вазочке.
Также из фоамирана можно сделать красивые цветы яблони.
Обьемная орхидея
Орхидея из белого фоамирана создается достаточно быстро, даже если вы новичок. Для этого нужно просто изучить наш урок и поэтапно следовать инструкции.
Чтобы создать необычный цветок, нам понадобится:
- Белый фоамиран, толщиной 1 мм.
- Стальная проволока 24-26 размера, вне зависимости от цвета.
- Ватные палочки и тейп-лента светло и темно-зеленого цвета.
- Сухая пастель, акриловая краска (оттенок фуксия) и цветные мелки.
- Тонкие кисти (лучше всего приобретите №0 или 1).
- Пинцет, маникюрные и канцелярские ножницы.
- Универсальный глиттерный клей, утюг и фактура орхидеи (Coerulea или Phalaenopsis 2 части).
Теперь можно начинать изготовление цветущих бутонов Фаленопсис, которое будет происходить в пять этапов.
- Создание шаблонов
Переносим на картон выкройку для цветов орхидеи двух размеров – S и M. На одну ветку нам понадобится 2-3 маленьких и 3-4 больших заготовки для бутонов, которые изображены на фото.
- Работа по вырезанию заготовок из фоамирана
Берем шаблон и переносим на фоамиран фигурный рисунок. Нам потребуется по одной заготовке для создания цветка: шаблон №1 (губа орхидеи), 2 (боковые лепестки или петалии), 3 (чашелистики – сепалии).
Шаблон № 5 — «кресло», оно же выступающая часть губы. Эту деталь вырезаем самостоятельно вручную. Нам потребуется сделать квадрат 6×6 мм, по краям которого нарезать зубчики с двух противоположных сторон, как показано тут.
Далее вырезаем по шаблону №4 бутон. На каждой ветке должно быть по 2-3 бутона, чтобы наш зефирный цветок смотрелся реалистично.
- Тонировка губ Фаленопсиса
Посредством сухих мелков и пористого аппликатора (губки) раскрашиваем заготовки в лиловый и ярко-желтый оттенок так, как показано на изображении.
Тонируем «кресло» нашего цветка, которое будет располагаться в центре. Для этого используем двусторонний метод окраски, применяя лиловый цвет.
При помощи тонкой кисти и краски создаем пятнышки-штрихи, которые характерны для орхидеи. Картинка показывает, в какой последовательности их наносить.
В результате должна получиться вот такая лицевая и обратная сторона губы.
Моделируем форму заготовок
Разогреваем подготовленный утюг в положении терморегулятора «шерсть». Прикладываем заготовки бутонов к наружной поверхности утюга, ждем пока материал нагреется и сам отстанет от поверхности. После этого он примет нужную форму.
Начинаем работу с шаблоном №2. Слегка придерживаем одну часть лепестка в руках, а вторую прикладываем к разогретой поверхности утюга. Пока материал не остыл, мелкий фоамиран прикладываем к молду и максимально плотно прижимаем его к пластиковой фактуре пальцами. Повторяем аналогичную процедуру со второй частью лепестка.
В итоге должны получиться объемные лепестки с прожилками, характерными для Фаленопсиса.
Оживляем наши лепестки, используя технологию вытягивания концов.
Формируем соответствующее углубление кончиками пальцев, чтобы получились вот такие лепестки.
Приступаем к работе с чашелистиками, используя для этого технику работы с шаблоном №2.
В результате должны получиться вот такие детали, которые показаны на фотографии.
Формируем губу, прикладывая лицевую сторону к утюгу. Материал должен нагреться и принять ту форму, которая нам необходима. Пока фоамиран еще теплый, создаем бороздку, защепив складку.
Приклеиваем «кресло» к сердцевине в то место, которое обозначено оранжевым кругом на первом шаблоне.
Подкрашиваем зубчики, используя желтый оттенок немного ярче «кресла».
- Сборка цветка
Формируем колонку путем нанесения клея на вату. Распределяем клей по вате. Если хотите получить бархатистую поверхность, то пока клей не застыл окуните палочку в крахмал.
Когда клей немного подсох, обрезаем головку ватной палочки, примерно 7-8 мм. Берем проволоку и на одном конце формируем маленькую петлю, которую промазываем клеем и продеваем вовнутрь головку ватной палочки. Фиксируем.
Приклеиваем к головке губу орхидеи, боковые лепестки и чашелистики.
Далее сборка бутонов:
Наша орхидея готова. С таким украшением любая свадебная прическа или резинка для волос станет максимально элегантной. А если вы «посадите» цветок в красивый горшок, то его никто не отличит от настоящего.
Еще один вариант создания орхидеи:
Предыдущее
Следующее
Предыдущее
Следующее
Мастер-класс «Изготовление розы из фоамирана» — Вектор-успеха.рф
Брошь — роза из фоамирана, украшение своими руками.
Материалы и инструменты
| Фоамиран тонкий в 1 миллиметр толщиной белого цвета длиной 25 сантиметров и шириной 25 сантиметров для лепестков | 18 страниц |
| Фоамиран зеленого цвета длиной 25 сантиметров и шириной 25 сантиметров для листьев и чашелистика | |
| Картон для шаблонов | |
| Готовая застежка для броши 3 сантиметра | |
| Фольга длиной 9 сантиметров и шириной 7 сантиметров для основы розы | |
| Ножницы средние для вырезания деталей цветов | |
| Клеющий пистолет для склеивания лепестков и листьев цветка | |
| Нижняя часть пресс-формы листа | |
| Утюг для обработки листьев и лепестков | |
| Зубочистка для обрисовки деталей шаблонов на фоамиране | |
| Маркер для обрисовки рисунка | |
| Проволока для стебля и листьев розы | |
| Масляная пастель для подкраски лепестков и листьев розы | |
| Тейп лента для обработки проволоки | |
| Линейка для разметки лент на фоамиране | |
| Кусачки для проволоки |
Потребуются инструменты и материалы:
Последовательность выполнения работы.
На фоамиране по линейке чертим спицей для вязания или зубочисткой полоску шириной 4 сантиметра.
\
Из полоски нарезаем квадратики 4 на 4 сантиметра.
Складываем квадратик по диагонали и вырезаем лепесток.
Можно вырезать сразу несколько лепестков по шаблону.
Лепестки вырезаны. Пышность розы зависит от количества лепестков. Примерное количество 12-20 лепестков
Окрашиваем лепестки у основания масляной пастелью светло зеленого и темно зеленого цвета.
Сначала окрашиваем светло зеленой пастелью.
Затем добавляем темно-зеленую пастель.
Лепестки окрашены
Нагреваем лепесток на утюге.
Руками подворачиваем оба края лепестка одновременно, вытягивая среднюю часть лепестка.
Вот такие лепестки у нас получились.
Делаем из фольги основу для розы в виде большой капли.
Приклеиваем вокруг заготовки для бутона первый лепесток с помощью клеевого пистолета.
Приклеиваем еще 2 лепестка первого ряда.
Приклеиваем 5 лепестков второго ряда.
Приклеиваем все лепестки третьего ряда.
Приклеиваем еще 4 лепестка, делая нашу розу более пушистой.
Нарезаем из зеленого фоамирана прямоугольники по размеру молда листа розы.
Фоамиран нагреваем на утюге и прикладывая к молду, делаем оттиск.
Оттиски для листочков готовы.
Квадратик из зеленого фоамирана складываем два раза и вырезаем чашелистик.
Вот такой чашелистик у нас получился. Можно вырезать чашелистик по шаблону.
Подкрашиваем масляной пастелью заготовки листьев и чашелистик.
Делаем надрезы по краям чашелистика и обрабатываем вручную, перетирая в пальцах рук.
Вырезаем листья.
Отрезаем проволоку и обрабатываем её тейп лентой.
К проволоке приклеиваем листья и собираем в трилистник.
Прикрепляем с помощью клеевого пистолета листья к розе.
Добавляем к броши два бутона.
С изнаночной стороны приклеиваем застежку для броши чашелистиком.
Брошь готова.
Фрагменты работы.
Спасибо за внимание.
Караева Tanya Karaeva Александровна (Tankar)
16.05.2018 г.
829
Чашелистный узор Жанны Лонг
Чашелистик — это лист, защищающий лепестки и внутреннюю часть распускающегося цветка. Наш Sepal — это аккуратные плечи с ажурной спинкой и широкой отделкой из кружевной резинки, которые можно носить вертикально или сложить в воротник-шаль. Это маленькая жемчужина свитера: идеально подходит для защиты и тепла, не жертвуя легкостью или стилем.
Конструкция: Sepal бесшовно сочетается с кардиганом реглан сверху вниз. Он начинается с предварительной наложения повязки и продвигается вперед и назад чуть ниже подмышек.Нежный узор из кружева, напоминающий цветок, проработанный по всей спине, создавая элегантный эффект взгляда. Ажурная ребристая кромка поднята и обработана круговыми швами на основной части, боковых сторонах и шее для получения бесшовной непрерывной отделки. На каждом из двух углов, где передние части встречаются с телом, несколько коротких рядов и стопка прибавок поворачивают углы и создают идеальную посадку.
Размер: Сепал сортируется по восьми размерам.
Fit: Sepal подходит для бюстов размером от 28 до 64 дюймов.Кружевная задняя часть Sepal довольно эластична, передняя часть спроектирована как , а не , чтобы перекрывать друг друга, а пожатие плечами выглядит довольно хорошо изношенным с несколькими дюймами положительного или отрицательного легкости. Советы по достижению наилучшего соответствия включены в примечания к выкройке.
Пряжа и ярд: От двух до трех мотков Why Knot Fibers ’Siren DK, нежно-сочной смеси кашемира в сердце, в цветовой гамме Ripple. Приблизительно 300 (330, 370, 400, 425, 450, 475, 500) ярдов завершат вашу кардио.Для немного менее структурированного образа мы рекомендуем Why Knot Fibers ’Seduction DK, мягкую и плавную смесь льна, шелка и альпаки.
Иглы: A размер 8 US (4,5 мм) длинная круговая спица 32 дюйма (для тела, как вперед-назад, так и по кругу) и иглы с двойным острием 8 US (4,5 мм) (для оребрение втулки), размер или , необходимый для получения калибра.
Условные обозначения: Четыре маркера петель, желательно два из которых отдельные, и игла для гобелена для ткачества на концах.
Инструкции: Шаблон включает как нарисованные, так и полностью написанные инструкции.
Используемые техники: предварительный набросок, вязание основного кружева, изнаночная петля через заднюю петлю, работа вперед и назад, работа по кругу и короткие ряды.
Спасибо: . По мнению CC, задиристая модель «вглядываться в середину и заставлять работать» и Сюзик, главный редактор-волонтер.
Как смоделировано: Последние три фотографии, предоставленные г-жойCabrera, покажите Sepal из альтернативной пряжи — Seduction DK, мягкой и плавной смеси льна, альпаки и шелка от Why Knot Fibers.
Sepal — обзор | Темы ScienceDirect
5.2.3.1 Потребление и торговля
Чаши растений руб. продаются и потребляются в сушеном виде в большинстве регионов мира, за заметными исключениями Малайзии и Вест-Индии, где они продаются и используются в свежем виде (Mounigan и Бадри, 2007). Они используются в традиционной кухне для приготовления горячих и холодных травяных настоев и других напитков, джемов, кондитерских изделий, салатов, гарниров и десертов (Mohamed et al., 2012; Boucher et al., 2014; Сид-Ортега и Герреро-Бельтран, 2015). Их экстракты в форме сиропа, порошка или концентрата, кроме того, широко используются в пищевой и фармацевтической промышленности Европы и Северной Америки в качестве натуральных красителей и ароматизаторов (Cisse et al., 2012; Da-Costa-Rocha et al. ., 2014; Сид-Ортега, Герреро-Бельтран, 2015).
Для удовлетворения растущего спроса на ruber calyces как на внутреннем, так и на экспортном рынках, в Китае, Таиланде, Малайзии, Филиппинах, Мексике, Ямайке, Вест-Индии, Соединенных Штатах Америки ( США), Судан, Сенегал, Мали, Чад, Нигерия, Нигер, Египет и Танзания (Christian and Jackson, 2009; Torres-Morán et al., 2011; Атта и др., 2013; Юнг и др., 2013; Патель, 2014; Тахир и др., 2017). Фактически, во всем мире идентифицировано более 100 культурных сортов или штаммов семян расы ruber (Plotto et al., 2004). Китай и Таиланд в настоящее время являются крупнейшими производителями этой культуры, контролируя значительную долю мировых поставок, в то время как Германия и США являются основными странами-импортерами (Da-Costa-Rocha et al., 2014).
Чашечки гибискуса продаются в основном сушеными, целыми и расфасованными в тюки, причем их качество сильно зависит от происхождения.Считается, что тайские сорта превосходят китайские, в то время как лучшие в мире культуры, как утверждается, поступают из Судана, крупнейшего африканского производителя и экспортера этого товара (Plotto et al., 2004; Mohamed et al., 2012). Тем не менее суданские чашечки немногочисленны, и их конечное качество часто ухудшается из-за плохой обработки (Mohamed et al., 2012). Между тем, Мексика, Ямайка, Египет, Сенегал, Танзания, Мали и Чад также становятся важными поставщиками. Но даже несмотря на то, что их производство за последние годы значительно улучшилось (Mc Clintock and El Tahir, 2004; Cisse et al., 2009a; Mohamed et al., 2012), основная его часть по-прежнему продается и потребляется внутри страны (Plotto et al., 2004).
Многие сорта (Американа, Тепалкатепек, Диаманте, Колима, Темпранильо, Талпа, Виолента, Судан и другие) выращиваются в основных районах производства гибискуса в Мексике (Torres-Morán et al., 2011; Borrás-Linares et al. , 2015). Между тем, четыре основных сорта гибискуса в настоящее время выращиваются в Сенегале для производства чашечек: суданский Vimto, местный Koor, тайский тайский и мексиканский CLT 92.Тайский и CLT 92 были недавно интродуцированы в этой стране, поэтому культивируются менее активно, чем традиционные сорта Вимто и Коор (Cisse et al., 2009a). Считается, что растения Vimto дают самые лучшие чашечки и самые высокие урожаи (до 5000 кг га, — 1 свежих чашек) (Mc Clintock and El Tahir, 2004; Plotto et al., 2004), поэтому выращиваются во всех сельскохозяйственных районах Западный Сенегал (Juliani et al., 2009). В этом регионе также широко культивируются растения коор, но их урожайность значительно ниже (2500–3000 кг га, –1 свежих чашечек) (Cisse et al., 2009а).
almaan / sepal: sepal: Пространственное выражение PAttern Locator
Это репо содержит:
- пакет sepal python и установочные файлы
- руководств и примеров использования (CLI и как импортированный пакет)
- данные, использованные в публикации
- результатов представлены в публикации
Загрузка и установка
sepal требует python3 , предпочтительно более позднюю или равную 3.5. Чтобы
загрузите и установите, откройте терминал и перейдите в каталог, в котором хотите
sepal для загрузки и выполнения:
git clone https://github.com/almaan/sepal.git cd чашелистик chmod + x setup.py ./setup.py установить
В зависимости от ваших прав пользователя вам может потребоваться добавить --user в качестве аргумента к setup.py .
Запуск установки даст вам минимально необходимую установку для расчета времени распространения. Тем не мение,
если вы хотите использовать модули анализа, вам также необходимо установить рекомендуемые пакеты.Для этого просто (в том же каталоге) запустите:
, возможно, потребуется включить --user . Кроме того, вам, возможно, придется использовать pip3 , если вы настроили интерфейс python-pip таким образом. Если вы используете conda или виртуальные среды, следуйте их рекомендациям по установке пакетов.
Это должно установить как интерфейс командной строки (CLI), так и стандартный пакет.
Чтобы проверить, была ли установка успешной, вы можете попробовать выполнить команду:
Который должен напечатать справочное сообщение, связанное с чашелистником.Если до сих пор у вас все получалось,
вы можете перейти к разделу примеров, чтобы увидеть sepal в действии!
Примеры
CLI
Чашелистник рекомендуется использовать
интерфейс командной строки. Оба моделирования
чтобы также вычислить времена диффузии
как последующий анализ или проверка результатов
можно легко выполнить, набрав чашелистик , а затем
либо запустить , либо проанализировать . Модуль анализа имеет разные
опции, чтобы визуализировать результаты ( осмотреть ),
сортировать профили по семействам паттернов ( семейство ) или по предметам
идентифицированные семьи к функциональному анализу обогащения ( fea ).Для
полный список доступных команд, выполните sepal module -h , где module
является одним из запускает и анализирует . Ниже мы проиллюстрируем
как чашелистик может быть использован для поиска профилей транскрипции с пространственными паттернами.
Мы создадим папку для хранения наших результатов, в которой также будет фигура
как наш рабочий каталог. Из основного каталога репо выполните:
cd res mkdir пример CD, пример
Образец MOB будет использован для иллюстрации нашего анализа.Мы начинаем
с расчетом времени распространения для каждого профиля транскрипции:
чашелистик запустить -c ../../data/real/mob.tsv.gz -mo 10 -mc 5 -o. -ар 1
Ниже пример (с дополнительным отображением команды справки)
как это может выглядеть
Вычислив времена диффузии, мы хотим проверить результат, например
В рамках исследования мы рассмотрим 20 лучших профилей. Мы легко можем сгенерировать
изображения из нашего результата, выполнив команду:
анализировать чашелистик -c../../data/real/mob.tsv.gz \ -r 20200409173043610345-top-diffusion-times.tsv \ -ар 1к -о. проверить -ng 20 -nc 5
Что будет выглядеть примерно так:
Результатом будет следующее изображение:
Затем, чтобы отсортировать 100 лучших генов по
набор семейств паттернов, где 85% дисперсии в наших паттернах
следует объяснять собственными паттернами, do:
sepal analysis -c ../../data/real/mob.tsv.gz \ -r 20200409173043610345-top-diffusion-times.tsv \ -ар 1к -о. семья -ng 100 -nbg 100 -eps 0,85 --plot -nc 3
Отсюда получаем следующие три
репрезентативных мотивов для каждой семьи:
Мы можем подвергнуть наши семьи дополнительному анализу, запустив:
sepal analysis -c ../../data/real/mob.tsv.gz \
-r 20200409173043610345-top-diffusion-times.tsv \
-ар 1к -о. fea -fl mob.tsv-family-index.tsv -or "mmusculus" , где мы, например, видим, что Семейство 2 обогащено несколькими процессами, связанными с функцией нейронов, генерацией и регуляцией:
| семья | родной | название | p_value | источник | размер_ пересечения | |
|---|---|---|---|---|---|---|
| 2 | 2 | GO: 0007399 | Развитие нервной системы | 0.00035977 | GO: BP | 26 |
| 3 | 2 | GO: 0050773 | регуляция развития дендритов | 0,000835883 | GO: BP | 8 |
| 4 | 2 | GO: 0048167 | регуляция синаптической пластичности | 0,00196494 | GO: BP | 8 |
| 5 | 2 | GO: 0016358 | Развитие дендритов | 0.00217167 | GO: BP | 9 |
| 6 | 2 | GO: 0048813 | морфогенез дендритов | 0,00741589 | GO: BP | 7 |
| 7 | 2 | GO: 0048814 | регуляция морфогенеза дендритов | 0,00800399 | GO: BP | 6 |
| 8 | 2 | GO: 0048666 | Развитие нейронов | 0.0114088 | GO: BP | 16 |
| 9 | 2 | GO: 0099004 | Путь передачи сигналов кальмодулин-зависимой киназы | 0,0159572 | GO: BP | 3 |
| 10 | 2 | GO: 0050804 | модуляция химической синаптической передачи | 0,0341913 | GO: BP | 10 |
| 11 | 2 | GO: 0099177 | регуляция транс-синаптической передачи сигналов | 0.0347783 | GO: BP | 10 |
Конечно, этот анализ ни в коем случае не является исчерпывающим. Но это довольно быстрый пример, показывающий, как использовать CLI для sepal .
Как импортная упаковка
Хотя sepal был разработан как отдельный инструмент, мы также создали его
функционировать как стандартный пакет Python, из которого могут быть
импортированы и используются в интегрированном рабочем процессе. Чтобы показать, как это можно сделать, мы
приведите пример, воспроизводящий анализ меланомы.Позже могут быть добавлены другие примеры.
Поддерживаемые файлы и формат
Требуется, чтобы входные данные для sepal были в формате n_locations x n_genes ,
однако, если ваши данные структурированы противоположным образом ( n_genes x n_locations )
просто укажите флаг --transpose при запуске симуляции или
анализ, и об этом мы позаботимся.
В настоящее время мы поддерживаем .csv , .tsv и .форматы h5ad . Для последнего ваш файл должен быть структурирован
в соответствии с ЭТОМ форматом. Мы ожидаем релиза от
scanpy Команда в ближайшем будущем, где будет представлен стандартизированный формат пространственных данных, но до тех пор мы будем
используя вышеупомянутый стандарт.
Данные
Все реальные данные, которые мы использовали, являются общедоступными, и их можно найти по следующим ссылкам:
- MOB: ССЫЛКА, используйте Rep11
- Мозг мыши: ССЫЛКА
- Лимфатический узел: ССЫЛКА
- Меланома: ССЫЛКА, используйте ST_mel1_rep1
- Мозжечок: ССЫЛКА, используйте Cerebellum_Puck_180819_11
Синтетические данные были сгенерированы:
- смешанный набор 1 (на основе изображений):
синтетический / img2cnt.py - смешанный набор 2 (паттерны Тьюринга):
synt / turing.py - наборы для абляции:
синтетический / ablation.py
Результаты
Все результаты, представленные в исследовании, можно найти в папке res , как для
реальные и синтетические данные. Для каждого образца мы структурировали результаты соответственно:
-
разрешение /-
название образца /-
X-времена диффузии.tsv: время распространения для всех ранжированных генов -
анализ /: содержит вывод вторичного анализа
-
-
Различные паттерны развития цветков в мутовчатых цветках, на примере Apiaceae и Brassicaceae в JSTOR
Abstract
Изучение последовательности инициации цветковых органов ценно не только для полимерных цветков, но и для мутовчатых цветков с фиксированным числом органов. Они показывают последовательность различных оборотов, а также последовательность органов в каждом обороте.В мутовчатых цветках эвдикотиледонов цветочная пластичность по количеству и расположению цветочных органов невелика. В этом исследовании будет показано, что ряд различных путей развития может приводить к относительно неизменной морфологии зрелых цветков. В цветках Apiaceae сохраняется одинаковое количество и положение цветковых органов, несмотря на значительные различия в раннем паттерне последовательности. Примечательны временные перекрытия между последующими оборотами, то есть перекрытие между инициациями различных типов органов.В цветках Brassicaceae единообразный филлотаксис цветков также обусловлен последовательностями, которые различаются в деталях. Реализованы почти все возможности в последовательности четырех лепестков, двух более коротких наружных тычинок и четырех более длинных внутренних тычинок (например, 4 лепестка → 2 тычинки → 4 тычинки или 2 тычинки → 4 лепестка → 4 тычинки или 4 лепестка → 4 тычинки → 2 тычинки и т. д.). Следующие морфогенетические выводы можно сделать из подробных исследований последовательности: (1) верхушка цветка может производить более одной категории органов одновременно.(2) Последовательность оборотов не должна быть строго акропетальной. (3) Положение органа и последовательность инициации органа, с одной стороны, и идентичность органа, с другой стороны, регулируются на разных генетических уровнях.
Информация о журнале
Текущие выпуски теперь размещены на веб-сайте Chicago Journals. Прочтите последний выпуск. С 1875 года Международный журнал наук о растениях (IJPS) представляет высококачественные, оригинальные, рецензируемые исследования лабораторий по всему миру во всех областях науки о растениях.Охватываемые темы варьируются от генетики и геномики, биологии развития и клеточной биологии, биохимии и физиологии до морфологии и анатомии, систематики, эволюции, палеоботаники, взаимодействия растений и микробов и экологии. IJPS приветствует статьи, в которых представлены оценки и новые взгляды на актуальные области биологии растений.
Информация об издателе
С момента своего основания в 1890 году в качестве одного из трех основных подразделений Чикагского университета, University of Chicago Press взяла на себя обязательство распространять стипендии высочайшего стандарта и публиковать серьезные работы, способствующие образованию, развитию общественное понимание и обогащение культурной жизни.Сегодня Отдел журналов издает более 70 журналов и сериалов в твердом переплете по широкому кругу академических дисциплин, включая социальные науки, гуманитарные науки, образование, биологические и медицинские науки, а также физические науки.
Изменчивость в контроле клеточного деления, лежащая в основе формирования эпидермального паттерна чашелистика Arabidopsis thaliana
Рисунок 3.
Модель интеркалярного роста воспроизводит распределение клеток по размерам.
(A, B) Живое изображение инициации зачатка чашелистника дикого типа, показывающее, что на основе анализа клонов внешний эпидермис чашелистника происходит примерно из двух рядов по 8 клеток (обведены белым) (видео S5). Эпидермальные ядра ( pATML1 :: h3B-mYFP ) окрашены в золото, а клеточные стенки (PI) — в зеленый цвет. Каждая клетка и все ее потомство помечены точкой одного цвета. Яркая точка, сидящая на вершине чашелистика на (B), — это пыльца. Масштабные линейки: 10 мкм. (C) Один кадр из живого изображения боковой чашелистики (обведен белым) (видео S6).Плазменные мембраны также отмечены золотом ( pATML1 :: mCitrine-RCI2A ). Обратите внимание, что верхняя часть чашелистника содержит дифференцированные замыкающие клетки (звездочка gc), и клетки больше не делятся, тогда как клетки в нижней части чашелистика активно делятся (дочерние клетки, произошедшие в течение последних 6 часов, обведены белым). Масштабная линейка: 50 мкм. (D) Диаграмма разброса, показывающая вертикальное положение каждого события деления (видео S6) в процентах от длины чашелистника (1 = вверху и 0 = внизу). Красная стрелка указывает на прогрессирующее базипетальное окончание делений.(E – F) Живое изображение более старого зачатка чашелистника, отображаемое каждые 12 часов (E: день 0) до зрелости на стадии 12 (F: 7,6 дня). Клетки из середины зачатка чашелистника и их потомство отслеживали по всей 7-й последовательности и использовали для определения областей зачатка и соответствующих областей зрелого чашелистика (обведены белым). Эти регионы произвольны и зависят только от ячеек, выбранных для отслеживания. Обратите внимание, что верхняя половина зачатка составляет только верхушку зрелого чашелистика, тогда как середина зачатка составляет верхнюю половину чашелистика, а несколько нижних слоев клеток составляют всю нижнюю половину зрелого чашелистика, что указывает на то, что эти нижние клетки продолжали размножаться.Масштабные линейки: 50 мкм. (G) Модель интеркалярного роста (IG). Вычислительная чашелистик развивается из упрощенного генеративного слоя из 8 клеток. В модели клетки генеративного слоя пролиферируют на протяжении всего развития. Верхнее потомство генеративного слоя входит в деления формирования паттерна и завершается после прохождения трех клеточных циклов, будь то митотические или эндоциклы. Последнее изображение справа имеет уменьшенное увеличение. Цвет клеток соответствует плоидности: белый, генеративный слой 2С; синий, 2С; зеленый, 4С; пурпурный, 8С; и красный, 16С.(H) Гистограмма, показывающая, что клетки продуцируются в четырех точных размерах с помощью модели IG, когда допускается изменение только эндоредупликации. Длина клеточного цикла постоянна, а деления точно симметричны (сравните с панелью K). Ось площади масштабируется с логарифмической базой 2. (I) Гистограмма времени клеточного цикла (с шагом 6 часов), измеренная на основе данных визуализации в реальном времени дикого типа (видео S1, S2, S3). (J) Гистограмма участков клеток в зрелом эпидермисе чашелистника, определенных полуавтоматической обработкой изображений (серый цвет; подробности см. В тексте S1).Ось площади масштабируется по логарифмическому основанию 2. Плоидность клеток откалибрована с помощью 47 ячеек дорожного покрытия, для которых как содержание ДНК, так и площадь известны из рисунка 1F (размер области подчеркнут). Обратите внимание, что площади ячеек имеют широкое распределение, хотя пики для 2C и 4C видны. (K) Гистограмма площадей клеток, полученных с помощью модели IG, включая изменчивость времени клеточного цикла и шум в симметрии деления, показывающая, что каждый уровень плоидности имеет распределение размеров клеток. Общее распределение по размерам существенно не отличается от распределения in vivo (рис. 3J) (см. Текст S1 для дальнейшего анализа).См. Также видео S4 – S6.
Подробнее »
БУТЫЛКИ ПОП ЗАВЕРШИТЬ ЦВЕТОЧНЫЙ ПРОЕКТ
Презентация на тему: «ПОП-БУТЫЛКА ЗАВЕРШИТЬ ЦВЕТОЧНЫЙ ПРОЕКТ» — стенограмма презентации:
1
ПОП-БУТЫЛКА ЗАВЕРШЕННЫЙ ЦВЕТОЧНЫЙ ПРОЕКТ
Представлено: Бобом Брауном
2
ПОЛНЫЙ ЦВЕТОК
3
Поп-бутылки Вам понадобятся прозрачная пластиковая бутылка на 1 литр и зеленая пластиковая бутылка на 1 литр той же марки.
4
Удаление этикеток Наполните бутылки горячей водой и оставьте на пару минут. Затем медленно удалите этикетку. Этот процесс удалит большую часть клея.
5
Удалите излишки клея. Используйте средство для удаления клея, которое не будет токсичным для учащихся. Я использовала средство для удаления клея на основе цитрусовых, которое смывается водой и не опасно.
6
Создание выкройки Плоская поверхность бутылки имеет размеры 10 дюймов на 3 ½ дюйма. Я использовал 8 лепестков или чашелистиков, чтобы узор представлял собой прямоугольники размером 1 1/4 дюйма на 3 1/2 дюйма.
7
Выкройка лепестка Теперь в каждом прямоугольнике вы должны нарисовать узор вашего лепестка. Помните, что основание лепестка должно оставаться прикрепленным к бутылке.
8
Рисунок чашелистика Рисунок чашелистика может быть таким же, или вы можете его изменить.
9
Перенос выкройки Вырежьте выкройки и прикрепите их к нужной бутылке. Лепестки на прозрачной бутылке и чашелистики на зеленой бутылке. Используйте перманентную ручку, чтобы отметить рисунки.
10
Вырезание узора из бутылки
Удалите воду, не стирайте следы узора.Используйте нож, чтобы открыть место на бутылке, куда можно вставить ножницы, чтобы вырезать узор.
11
Раскрасьте лепестки. Используйте перманентный маркер, чтобы раскрасить лепестки прозрачной бутылки. Раскрасьте обе стороны для более глубокого цветового рисунка. Не окрашивайте остальную часть бутылки. Отрежьте прозрачное горлышко бутылки.
12
Пестик Вы готовы сделать части пестика; клеймо, стиль и завязь.Клеймо — это маленькая бусинка, в виде цветного ершика, а на завязи — помпон.
13
Тычинки Тычинки представляют собой цветной шарик для пыльника и средство для очистки трубок для нити. Вы должны сделать четыре штуки, чтобы они выглядели одинаково.
14
Репродуктивная сборка
Нанесите большое количество горячего клея на центр зеленой крышки бутылки.Поместите четыре тычинки в виде + в центр шляпки. Поместите пестик поверх них. Держите, пока не закрепите.
15
Полные части цветка
Вы изготовили все детали, необходимые для сборки вашего цветка.
16
Сборка Возьмите прозрачную бутыль с лепестками и вставьте ее внутрь зеленой бутылки с чашелистиками.Выровняйте их так, как вы хотите, а затем соедините их вместе. Наденьте колпачок.
17
Готовый цветок Это вид сбоку вашего целого цветка.
18
Готовый цветок Это вид сверху вашего целого цветка.
Клоны клеток переходят от уменьшения к увеличению изменчивости размеров чашелистиков Arabidopsis | Развитие
В процессе развития органы формируются с удивительно постоянными размерами и формой, тогда как на клеточном уровне наблюдается высокая вариабельность роста.Учитывая этот контраст, неясно, как такая согласованность в масштабе органов может возникнуть из-за клеточного поведения. Здесь мы исследуем промежуточную шкалу роста клонов клеток в чашелистике Arabidopsis . Каждый клон состоит из потомства одной клетки-предшественника. На ранних стадиях мы обнаружили, что клоны, полученные из маленькой клетки-предшественника, растут быстрее, чем клоны, полученные из большой клетки-предшественника. Это приводит к уменьшению изменчивости размеров клонов, явление, которое мы называем униформизацией размера.Напротив, на более поздних стадиях роста клонов клоны изменяют свой характер роста для увеличения изменчивости размеров, когда клоны, полученные из более крупных клеток-предшественников, растут быстрее, чем клоны, полученные из более мелких клеток-предшественников. Наконец, мы обнаружили, что на ранних стадиях быстрорастущие клоны демонстрируют большую гетерогенность клеточного роста. Таким образом, клеточная изменчивость в росте может способствовать снижению изменчивости клонов по всему чашелистику.
Как и у большинства живых многоклеточных организмов, органы растений воспроизводимы; органы имеют свои собственные характерные размеры и формы, что делает их ориентирами для определения видов в ботанике.Наивно, мы могли бы ожидать, что воспроизводимые органы возникнут из однородных клеток с постоянным размером и формой, как плитка на полу. Однако визуализация в реальном времени демонстрирует, что во многих случаях размеры, темпы роста и направления клеток демонстрируют значительную изменчивость (Roeder et al., 2010, 2012; Hong et al., 2016; Elsner et al., 2012; Kierzkowski et al., 2012 ; Tauriello et al., 2015; Uyttewaal et al., 2012). Например, время и геометрия деления клеток различны в органах стереотипной формы (Roeder et al., 2010; Бессон и Дюмэ, 2011). Эти результаты поднимают вопрос о вкладе такого клеточного шума и изменчивости в согласованность размера / формы органов (Meyer and Roeder, 2014; Hong et al., 2016).
Сложность взаимоотношений между клетками и органами можно проиллюстрировать несколькими известными и загадочными биологическими примерами. Первый пример — «регулирующее яйцо»; на определенных ранних стадиях, когда половина раннего эмбриона Xenopus удаляется, оставшаяся половина дает головастика половинного размера (Spemann and Mangold, 1924; Cooke, 1975).Это предполагает, что судьба клеток может определяться относительным расположением внутри эмбриона. В этом сценарии клетки не будут полностью автономными, а будут подчиняться всей форме и функциям эмбриона. Второй пример — «компенсация»; когда мутация подавляет деление клеток и, следовательно, уменьшает количество клеток в органе, и отдельные клетки компенсируют эту потерю, увеличивая свой размер, чтобы произвести орган почти правильного размера и формы (Tsukaya, 2003). Этот феномен компенсации предполагает, что у органов есть глобальный механизм определения размера / формы, который делает рост клеток подчиненным размеру / форме всего органа.Тем не менее, как упоминалось выше, клетки сохраняют способность демонстрировать переменную скорость роста, что предполагает, что клетки также в значительной степени автономны (Asl et al., 2011; Elsner et al., 2012). Таким образом, мы остаемся с картиной, в которой развитие является результатом баланса между организменной теорией (Kaplan and Hagemann, 1991; поведение клеток является следствием поведения органов) и теорией клеток (поведение органов является следствием поведения клеток). . Чтобы пролить свет на механизмы, уравновешивающие индивидуальное и коллективное поведение при росте клеток, мы решили сосредоточиться на промежуточном масштабе, группах клеток, используя кинематический подход.
Здесь мы сосредотачиваемся на клоне (т. Е. Группе родственных клеток, которые происходят от одной клетки-предшественника) в чашелистике Arabidopsis как попытке идентифицировать объединяющий механизм, который также может быть совместим как с клеточной теорией, так и с организменным. теория. Интересно, что Tauriello et al. (2015) использовали кинематический подход для извлечения роста клонов, чтобы определить общие свойства кривых роста.К удивлению, они обнаружили, что размеры разных клонов следуют одной и той же сигмоидальной функции времени, хотя и со стохастической синхронизацией максимальной скорости роста, подразумевая, что клоны не растут «свободно», а вместо этого ограничены. Поскольку эти кривые роста начинаются с разных начальных размеров клеток, точный вклад начального распределения по размерам в такие модели роста становится центральным вопросом. В этом исследовании мы исследовали подробную кинематику и взаимосвязь между поведением роста и стартовыми размерами клонов у чашелистиков Arabidopsis и .
Во-первых, мы исследовали взаимосвязь между начальными размерами клонов и скоростью их роста в развивающихся чашелистиках Arabidopsis . Здесь клон относится к клетке-предшественнику и всем ее потомкам, и в дальнейшем мы будем использовать «изначально маленький (или большой) клон» для клона, произошедшего от маленькой (или большой) клетки-предшественника. Мы проверили, стали ли размеры клонов внутри чашелистика более однородными (унификация размеров) или более изменчивыми (увеличение вариабельности размеров) с течением времени.Данные визуализации в реальном времени из двух лабораторий (пять чашелистиков дикого типа), о которых ранее сообщалось в Hervieux et al. (2016). В этом исследовании клетки были очерчены маркерами плазматической мембраны, а изображение всего чашелистика отображалось каждые 12 или 24 часа. Мы рассматривали рост всего клона как единицы и игнорировали деления клеток внутри клона. Рост отдельных клеток будет обсуждаться в разделе, озаглавленном «Гетерогенность роста отдельных клеток положительно коррелирует с ростом клонов на каждом временном шаге».Чтобы извлечь схему и проследить за ростом клонов, мы использовали программное обеспечение для анализа и визуализации MorphoGraphX (MGX) (Barbier de Reuille et al., 2015; см. Материалы и методы) для сегментации клеток, отслеживания клонов и расчета площади. Мы определили площадь клона в момент времени t как A t и рассчитали относительный рост клона как ( A t + Δ t — A t ) / A t × 100 (%) (рис.1). Характер роста у всех проанализированных чашелистиков был качественно подобен (рис. S1A-D).
Рис. 1.
Пространственно-временная картина роста клонов. Тепловая карта ареального роста клонов ( A т + Δ т — A т ) / A т × 100 (%) в течение последовательных 12 часов интервалы для цветка wt-a1.Клоны обведены черным контуром. Новые клеточные стенки, полученные в результате делений внутри клона, отмечены белым цветом. Обратите внимание, что на стадии 5-7 цветков наблюдается более высокая скорость роста в ночное время и относительно более низкая скорость роста в течение дня. Масштабная линейка: 50 мкм.
Рис. 1.
Пространственно-временная картина роста клонов. Тепловая карта ареального роста клонов ( A т + Δ т — A т ) / A т × 100 (%) в течение последовательных 12 часов интервалы для цветка wt-a1.Клоны обведены черным контуром. Новые клеточные стенки, полученные в результате делений внутри клона, отмечены белым цветом. Обратите внимание, что на стадии 5-7 цветков наблюдается более высокая скорость роста в ночное время и относительно более низкая скорость роста в течение дня. Масштабная линейка: 50 мкм.
Наше живое изображение чашелистиков начинается вскоре после того, как зачатки чашелистиков сформировались на боках цветочной меристемы, когда клетки относительно малы по размеру и еще не дифференцировались в специализированные типы эпидермальных клеток.Как сообщалось в нашем предыдущем исследовании (Hervieux et al., 2016), чашелистик сначала подвергается быстрому росту на кончике (стадия 3-4 на рис. 1). Затем максимальный рост постепенно перемещается вниз к середине (стадия 5-7) и низу чашелистика (стадия 8-9) на рис. 1 (см. Также рис. S1A-D). Общая тенденция роста площади клонов в каждом чашелистике может быть отражена в среднем A t (фиг. 2A). Хотя чашелистики из разных лабораторий (wt-a1, wt-a2, wt-b1, wt-b2 и wt-b3 на рис.2А) немного отличаются из-за разных условий культивирования растений, чашелистики в данной лаборатории демонстрируют сопоставимые кривые роста. На рис. 2 и в таблице 1 мы представляем стадии цветения чашелистиков, определенные на основе Smyth et al. (1990) (см. Также «Материалы и методы»).
Рис. 2.
Унифицированность размеров и повышение изменчивости клонов. (A) Средняя площадь клонов с течением времени.(B) CV для количественной оценки изменчивости площади. Планки погрешностей представляют собой доверительный интервал 50% (см. Материалы и методы). CV сначала уменьшается, а затем увеличивается. Обратите внимание на то, что wt-a1 чашелистика не показывает очевидного увеличения, что может быть связано с тем, что исходные клетки wt-a1 отображались в основном от кончика до средней части чашелистника. (C) Схематическое изображение трех тенденций роста с более ранним размером клона (пунктирные линии) и более поздним размером клона (сплошные линии): унифомизация размера (A1), первоначально меньшие клоны растут быстрее; эквивалентный рост (A2), клоны растут независимо от их первоначальных размеров; повышение вариабельности (A3), изначально более крупные клоны растут быстрее.(D, E) в зависимости от графика для клонов (D) и графика (E). Пунктирные линии представляют собой линейные аппроксимации точек данных на графике зависимости от . (F) Примеры различных тенденций роста, показывающие линейную корреляцию между областями в более позднее время и в более раннее время . Кривые 1 (зеленый) и 3 (розовый) иллюстрируют униформизацию, кривая 2 (синяя) показывает эквивалентный рост, а кривая 4 (красная) показывает увеличение изменчивости. (G, H) по сравнению с графиком для клонов при T 0 по сравнению с клонами при T 1 (G) и при T 1 по сравнению с T 2 (H) .Пунктирные линии в G и H соответствуют точкам данных, соответствующим точкам D и E, соответственно. (I) Примеры различных тенденций роста между и . Цвета линий такие же, как и в F. Унифицированность размера (увеличение изменчивости) соответствует случаю с отрицательной (положительной) корреляцией.
Рис. 2.
Унифицированность размеров и повышение изменчивости клонов. (A) Средняя площадь клонов с течением времени. (B) CV для количественной оценки изменчивости площади. Планки погрешностей представляют собой доверительный интервал 50% (см. Материалы и методы).CV сначала уменьшается, а затем увеличивается. Обратите внимание на то, что wt-a1 чашелистика не показывает очевидного увеличения, что может быть связано с тем, что исходные клетки wt-a1 отображались в основном от кончика до средней части чашелистника. (C) Схематическое изображение трех тенденций роста с более ранним размером клона (пунктирные линии) и более поздним размером клона (сплошные линии): унифомизация размера (A1), первоначально меньшие клоны растут быстрее; эквивалентный рост (A2), клоны растут независимо от их первоначальных размеров; повышение вариабельности (A3), изначально более крупные клоны растут быстрее.(D, E) в зависимости от графика для клонов (D) и графика (E). Пунктирные линии представляют собой линейные аппроксимации точек данных на графике зависимости от . (F) Примеры различных тенденций роста, показывающие линейную корреляцию между областями в более позднее время и в более раннее время . Кривые 1 (зеленый) и 3 (розовый) иллюстрируют униформизацию, кривая 2 (синяя) показывает эквивалентный рост, а кривая 4 (красная) показывает увеличение изменчивости. (G, H) по сравнению с графиком для клонов при T 0 по сравнению с клонами при T 1 (G) и при T 1 по сравнению с T 2 (H) .Пунктирные линии в G и H соответствуют точкам данных, соответствующим точкам D и E, соответственно. (I) Примеры различных тенденций роста между и . Цвета линий такие же, как и в F. Унифицированность размера (увеличение изменчивости) соответствует случаю с отрицательной (положительной) корреляцией.
Затем мы проанализировали, как изменчивость размера клона изменяется по мере развития чашелистика.Для этого мы рассчитали коэффициент вариации (CV, определяемый как стандартное отклонение, деленное на среднее значение) A t как функцию от t (рис. 2B). Существует три возможных тенденции в изменчивости площадей клонов, количественно оцененной с помощью CV (рис. 2C): (A1) униформализация размера, во время которой первоначально меньшие клоны растут быстрее, чтобы догнать более крупные клоны (верхняя панель), что приводит к снижение CV; (A2) эквивалентный рост, во время которого клоны растут гомогенно и независимо от их первоначальных размеров (средняя панель), что приводит к неизменному CV; (A3) усиление вариабельности, во время которого первоначально более крупные клоны растут быстрее, опережая меньшие клоны, что приводит к увеличению CV (нижняя панель).Мы обнаружили, что первоначально CV для каждого цветка снижается, что указывает на то, что клоны подвергаются униформизации по размеру. Затем рост клонов претерпевает стереотипный переход, после которого CV увеличивается, указывая на то, что клоны переключаются на усиление вариабельности размера. Таким образом, мы обнаружили, что существует переломный момент, когда клоны совершают переход от унификации размера к увеличению вариабельности размера во время развития чашелистника.
Чтобы проверить переход от унификации размеров к увеличению вариабельности, мы затем определили, как площадь каждого клона, измеренная на ранней стадии, соотносится с его площадью в более позднее время.Мы выполнили это сравнение как на этапе унификации размеров, так и на этапе повышения изменчивости. Для этого в нашем анализе мы сначала определили несколько релевантных моментов времени: T 0 , T 1 и T 2 . T 0 обозначает ранний период нашей визуализации в реальном времени между стадией 3 и 6 (Таблицы 1 и 2). T 1 обозначает время, когда CV достигает своего локального минимума, то есть точки перелома. T 2 обозначает более поздний момент времени после T 1 , который обычно находится около стадии 8 цветения (таблицы 1 и 2).Мы можем измерить совокупный коэффициент роста, разделив конечный размер клона на размер исходного клона. Для унификации размера мы ожидали, что изначально маленькие клоны будут иметь более высокие коэффициенты совокупного роста, чем изначально большие клоны [т.е. отрицательная корреляция между и , как показано на гипотетическом графике рис. 2I (кривые 1 и 3)]. Как и ожидалось, отрицательная корреляция между и наблюдалась в реальных данных из [ T 0 , T 1 ], подтверждая, что унификация размеров происходит до критической точки (рис.2G). Напротив, для увеличения изменчивости размеров мы ожидали, что изначально маленькие клоны будут иметь более низкие коэффициенты совокупного роста, чем изначально большие клоны [т.е. положительная корреляция между и , как показано на гипотетическом графике рис. 2I (кривая 4)]. Эта положительная корреляция между и наблюдалась после критической точки T 1 в реальных данных, подтверждая, что происходит увеличение изменчивости размеров.
Таблица 2.
Результаты подгонки для линейной зависимости за период [ T E , T L ] = [ T 0 , T 1 ] и период [ T E , T L ] = [ T 1 , T 2 ]
Затем мы дополнительно исследовали математическую взаимосвязь между площадью каждого клона, измеренной в ранний период времени, и площадью в более позднее время, чтобы найти ключевой параметр, отличающий униформизацию размера от режимов роста увеличения вариабельности размера.Во время периода унификации размера (A1), [ T 0 , T 1 ], мы наблюдаем линейную корреляцию между площадью клона на T 0 () и площадью клона на T 1 () (Рис. 2D). Мы наблюдаем различную линейную корреляцию между площадью клона при T 1 () и площадью клона при T 2 () во время периода увеличения вариабельности (A3) [ T 1 , T 2 ] (рис.2E). Более того, феноменологическая модель, которая подтвердила, что лежащие в основе сигмоидальные кривые роста этих клонов приводят к этим эмпирически наблюдаемым линейным отношениям, представлена в материалах и методах (см. Также рис. S2). Поскольку мы наблюдали различные линейные корреляции в и (рис. 2D, E), мы ожидали, что ключевой параметр для перехода между униформизацией размеров и повышением вариабельности может быть встроен в линейное уравнение, связывающее площадь клона в более раннее время () с клонировать область в более позднее время (), т.е.е. (Рис. 2F). Все клоны увеличивают свои площади с T E до T L ; следовательно, параметр α больше или равен 1. В этой схеме кумулятивный коэффициент роста (т.е. конечный размер клона, деленный на исходный размер клона), определяемый как, можно легко получить как. Как показано на гипотетических графиках (рис. 2F, I), случай унификации размеров (A1) возникает, когда β > 0 (например, кривые 1 и 3). Случай эквивалентного роста (А2) (рост клона не зависит от размера клона, т.е.е. не зависит от ) соответствует β = 0 (кривая 2). Случай увеличения изменчивости (A3) (первоначально более крупные клоны растут быстрее, т.е. положительно коррелирует с ) возникает, когда β <0 (например, кривая 4).
Подгоняемые параметры до точки перелома ( α 1 , β 1 ) удовлетворяют требованиям α 1 > 0 и β 1 > 0 для унификации (таблица 2).С другой стороны, после точки перелома ( α 2 , β 2 ) удовлетворяют α 2 > 0 и β 2 <0 для чашелистиков wt-a2, wt -b1, wt-b2 и wt-b3, что соответствует увеличению изменчивости. Однако β 2 > 0 для чашелистника wt-a1, потому что wt-a1 показывает более или менее A1 или A2 (рис. S3D), что согласуется с CV-анализом, показывающим, что wt-a1 остается в униформизированном состоянии и не достигает переломный момент (рис.2Б). Подводя итог, мы пришли к выводу, что во время развития чашелистника механизм униформизации размера снижает изменчивость площади клона на ранних этапах до перехода к другому поведению в переломный момент, после чего повышение изменчивости приводит к последующему увеличению изменчивости площади клона. Кроме того, мы обнаруживаем, что параметр β , соответствующий пересечению y на графике зависимости (рис. 2F), обеспечивает эффективный параметр для идентификации такого перехода роста.
Одно из следствий линейной положительной корреляции между более ранними размерами клонов и более поздними как в униформизации размера, так и в повышении вариабельности (Рис. 2D, E) состоит в том, что ранг размеров клона в значительной степени сохраняется во время развития чашелистика. Это означает, что, хотя более мелкие клоны растут быстрее на стадиях унификации размера, их темпы роста недостаточно высоки, чтобы догнать или даже обогнать изначально более крупные клоны.Мы задавались вопросом, сыграла ли позиционная информация роль в упорядочивании размера клона. Чтобы проверить это, мы отслеживали площади клонов A t как в пространстве, так и во времени на стадиях унификации размеров (рис. 3A, B), раскрашивая кривые роста в A t согласно их первоначальному размеру на момент времени T 0 (Рис. 3B). Цветовой градиент в более ранние времена в значительной степени сохраняется в более поздние времена (Рис. 3B; Рис.S4), еще раз подтверждая, что размерный порядок клонов в значительной степени сохраняется. В начальный момент времени T 0 меньшие клетки распределяются по направлению к верхней части чашелистника, а более крупные — к нижней. Это связано с тем, что клетки в верхней части находятся дальше в своем развитии и начали делиться на ранних стадиях цветения. Как и ожидалось, на пространственное распределение размеров рост не сильно влияет, и клетки на кончике остаются меньше, чем клетки на дне чашелистника в более позднее время T 1 (рис.3С; Рис. S4).
Рис. 3.
Рис. 3.
Затем мы рассмотрели пространственное распределение коэффициента совокупного роста на чашелистике (рис. 3D). Для всех проанализированных чашелистиков наблюдается непрерывное изменение от высоких значений в верхней части к более низким значениям в нижней части. Чтобы выяснить, происходит ли униформализация размеров только в определенных регионах (например,грамм. верхушку чашелистика, мы вручную разделили чашелистик на верхнюю, среднюю и нижнюю области, как показано голубым, синим и пурпурным цветами соответственно (рис. 3E; рис. S5). отрицательно коррелировал с для клонов из каждой области, показывая, что унификация размеров происходит по всему чашелистику (фиг. 3F; фиг. S5). Графики из разных регионов были немного смещены относительно друг друга, как и ожидалось, учитывая общие тенденции роста чашелистника; а именно, верхние клоны имеют меньший, но больший , тогда как нижние клоны имеют больший, но меньший размер (см. также рис.3G, H). Хотя распределения и немного различаются, как схематично показано на рис. 3I, наши результаты показывают, что униформизация размера происходит как глобальный механизм в развивающейся чашелистике, а информация о положении не играет значительной роли в регулировании размера клонов во время этапы унификации размеров. На этапах увеличения изменчивости мы не смогли найти характерного тренда (рис. S6).
Поскольку униформализация размеров не зависит от расположения клонов, мы затем спросили, есть ли постепенное или внезапное изменение во времени корреляции между начальным размером клона и коэффициентом роста ( A t + Δ t / A т ) / Δ т .Другими словами, мы исследовали корреляцию на коротких временных интервалах, в которых Δ t составляет 12 или 24 часа (по сравнению с более длинными временными интервалами, проанализированными в предыдущих разделах). Начальные размеры клонов отрицательно коррелируют с коэффициентами роста в течение первых 0-72 часов (или стадии 4-7), тогда как на более поздних стадиях корреляции становятся положительными (рис. 4A). Изменение корреляций с отрицательных на положительные было дополнительно подтверждено путем расчета на каждом временном шаге коэффициента корреляции Спирмена (SCC) между начальными размерами и коэффициентами роста ( A t + Δ t / A т ) / Δ т от всех клонов.SCC указывают на то, что коэффициенты роста на каждом временном шаге отрицательно коррелируют на более ранних стадиях, но становятся положительно коррелированными на более поздних стадиях (рис. 4B). Переход произошел примерно на стадии 6-7, за исключением того, что SCC для чашелистника wt-a1 имеет задержку по времени около 36 часов. Эти результаты также показывают, что существует временная вариация, изменяющаяся от отрицательной корреляции во время унификации размеров до положительной корреляции на стадиях поощрения изменчивости, как показано на рис.4B, что означает, что переход происходит постепенно.
Рис. 4.
Унификация размера зависит от времени запуска клона. (A) График коэффициента роста на каждом временном шаге ( A t + Δ t / A t ) / Δ t в зависимости от нормализованной начальной площади для вес- а2. Здесь рассматривается нормализованная начальная область для лучшего визуального сравнения областей в разные моменты времени.Делящиеся и неделящиеся клоны окрашены в красный и синий цвета соответственно. (B) SCC для графиков в A. SCC меняются с отрицательного на положительный примерно на стадии 6-7 (72-96 ч). (C) Схематическое изображение определения клона с момента начала T S . Пунктирные рамки показывают некоторые примеры определения ( T E = 12 часов для T S = 0 или T E = 24 часа для T S = 24).(D) Размер и скорость роста различных клонов. (E) SCC для различных комбинаций T E и T S .
Рис. 4.
Унификация размера зависит от времени запуска клона. (A) График коэффициента роста на каждом временном шаге ( A t + Δ t / A t ) / Δ t в зависимости от нормализованной начальной площади для вес- а2.Здесь рассматривается нормализованная начальная область для лучшего визуального сравнения областей в разные моменты времени. Делящиеся и неделящиеся клоны окрашены в красный и синий цвета соответственно. (B) SCC для графиков в A. SCC меняются с отрицательного на положительный примерно на стадии 6-7 (72-96 ч). (C) Схематическое изображение определения клона с момента начала T S . Пунктирные рамки показывают некоторые примеры определения ( T E = 12 часов для T S = 0 или T E = 24 часа для T S = 24).(D) Размер и скорость роста различных клонов. (E) SCC для различных комбинаций T E и T S .
Остается вопрос, связан ли переход от унификации размеров к увеличению вариабельности со сроками развития всего чашелистика или со временем, когда клон инициируется, поскольку наш анализ в предыдущих разделах начался с единичных клеток в первый раз. точка серии изображений в реальном времени.Чтобы ответить на этот вопрос, мы рассмотрели клоны, начинающиеся с отдельных клеток в дополнительные моменты времени во время последовательности визуализации в реальном времени данного чашелистика, обозначенное как начальное время T S , в которое извлекается набор всех отдельных клеток. с их клеточными стенками и идентифицированными участками (рис. 4C, крайние левые контуры). Каждая отдельная клетка, определенная как T S , поэтому служит клеткой-предшественником конкретного клона для t > T S , т.е.е. другой выбор T S (= 0 ч, 12 ч, 24 ч,…) приводит к другому выбору клонов (рис. 4C). Другой выбор T S эквивалентен разному времени начала (обозначенному как t = 0 ч в таблице 1) визуализации в реальном времени, которое может быть произвольным. Затем мы спросили, зависит ли признак униформизации размеров, т.е. отрицательные корреляции, наблюдаемые на рис. 4A, B, от выбора T S .
Как было выполнено ранее (рис. 4A), мы исследовали, как размер клона при t = T E (с T E > T S ) коррелирует с коэффициентом роста (с Δ t = 12 ч) на каждом временном шаге (например, прямоугольники с пунктирной линией на фиг. 4C для различных вариантов выбора T E ).На рис. 4D представлено несколько примеров зависимости от с различными вариантами T S и T E . Мы видим, что всякий раз, когда новый клон инициируется из одной клетки, он подвергается униформизации по размеру, т.е. существует отрицательная корреляция между размером клона и скоростью роста, что указывает на то, что меньшие клоны растут быстрее, чем более крупные клоны (рис. 4D). Одновременно клон, инициированный ранее, T S , мог уже перейти в режим повышения вариабельности.Например, клон, инициированный в 108 ч, подвергается униформизации по размеру, тогда как клон, инициированный в 0 ч, подвергается усилению вариабельности (крайняя правая панель на фиг. 4D). Поскольку единичные клетки, инициирующие клоны через 108 часов, включены в 0-часовые клоны, это составляет сложную многомасштабную систему.
Мы проверили, что этот переход относится к времени начала клона с SCC для разных T S и T E , в которых синие и красные области соответствуют униформизации размера (A1) и увеличения вариабельности (A3) соответственно (рис.4E; Рис. S7). Интересно, что переход роста между A1 и A3 представлен как T S . Кроме того, время перехода от синей к красной области на фиг. 4E — это период, в течение которого соответствующие SCC показывают, что переход роста от униформизации к повышению вариабельности постепенно смещается к более позднему времени в зависимости от начального времени T S . В среднем переход происходит через ~ 60 часов для дикого типа a (wt-a2) [~ 24 часа для дикого типа b (wt-b1, wt-b2 и wt-b3) на рис.S7] после инициации клона из одиночной клетки на T S . Это означает, что переход роста, наблюдаемый на фиг. 4A и B, зависит от возраста клона, т.е. клоны должны вырасти до определенного размера (или масштаба), чтобы произошло усиление изменчивости. Между прочим, это показывает, что переход не зависит от общего времени развития чашелистника.
Поскольку причина переломного момента не может быть связана просто с изменением идентичности клеток, мы затем исследовали, может ли гетерогенность роста внутри клона быть связана с переходом от униформизации к усилению вариабельности.Чтобы ответить на этот вопрос, мы количественно определили степень неоднородности роста клеток G h в момент времени t , которая определяется как CV коэффициента роста всех отдельных клеток в пределах конкретного клона (см. Материалы и Методы). Чтобы определить роль неоднородности роста клеток в униформизации размеров, мы сначала спросили, коррелирует ли G h с начальным размером (рис. 5A, B). Мы обнаружили, что корреляция между гетерогенностью роста и размером исходного клона в основном отрицательная на ранних стадиях и становится положительной на более поздних стадиях.Другими словами, более высокий уровень гетерогенности роста клеток существует среди дочерних клеток для первоначально меньших клонов на стадиях униформизации размера, в то время как на стадиях увеличения вариабельности первоначально более крупные клоны демонстрируют более высокую гетерогенность роста (рис. 5E).
Рис. 5.
Увеличение неоднородности клеточного роста и корреляции с исходным размером клона. (A) Гетерогенность роста клеток по сравнению с нормализованной начальной площадью клонов.(B) SCC A. (C) Гетерогенность роста клеток в зависимости от коэффициента роста клона на каждом временном шаге. (D) SCC C. Обратите внимание, что вес-a1 чашелистика имеет меньшую корреляцию на более поздних стадиях, что может быть связано с временной задержкой перехода к росту. (E) Схематическое изображение результатов для более раннего размера клона (пунктирные линии) и более позднего размера клона (сплошные линии).
Рис. 5.
Увеличение неоднородности клеточного роста и корреляции с исходным размером клона. (A) Гетерогенность роста клеток по сравнению с нормализованной начальной площадью клонов.(B) SCC A. (C) Гетерогенность роста клеток в зависимости от коэффициента роста клона на каждом временном шаге. (D) SCC C. Обратите внимание, что вес-a1 чашелистика имеет меньшую корреляцию на более поздних стадиях, что может быть связано с временной задержкой перехода к росту. (E) Схематическое изображение результатов для более раннего размера клона (пунктирные линии) и более позднего размера клона (сплошные линии).
Исходя из временного изменения корреляции между исходным размером и G h (рис.5B) и аналогичной между начальным размером и коэффициентом роста на каждом временном шаге (рис. 4B), ожидается положительная корреляция между G h и коэффициентами роста клонов на каждом временном шаге. как для стадий унификации размеров, так и для стадий увеличения изменчивости. Такие положительные корреляции были подтверждены на рис. 5C и D; а именно, более быстрорастущий клон на временном шаге Δ t (т.е. 12 ч в нашем случае) имеет большую гетерогенность роста (или вариабельность) среди отдельных клеток, принадлежащих клону.Результаты для других чашелистиков качественно аналогичны (рис. S9). Эти данные свидетельствуют о том, что изменчивость роста на клеточном уровне может способствовать росту клона. В качестве альтернативы, быстрорастущие клоны могут демонстрировать большую гетерогенность в скорости роста отдельных клеток, потому что они должны приспосабливаться к более медленному росту соседних клонов. Этот фактор может играть важную роль в растительных клетках, потому что клетки приклеены к своим соседям и не могут скользить друг по другу, как это делают клетки животных.Наконец, быстрорастущие клоны с большей вероятностью инициируют устьица, которые вначале растут быстрее, чем их ближайшие соседи, и поэтому могут вносить вклад в повышенную гетерогенность роста внутри клона.
В целом, эти анализы демонстрируют, что переход от унификации размеров к увеличению вариабельности происходит у растущих чашелистиков, что коррелирует с изменением неоднородности роста, а не с положением или идентичностью клеток (рис.5E).
Подводя итог, наши результаты показывают существование фазового перехода роста между двумя режимами роста в развивающейся чашелистике. Режим унификации размера происходит после инициации клонов, во время которого первоначально более мелкие клоны растут быстрее по сравнению с более крупными, что приводит к уменьшению вариабельности площадей клонов. Режим увеличения вариабельности происходит на более поздних стадиях клонов, во время которых первоначально более крупные клоны растут еще быстрее, что приводит к увеличению вариабельности площадей.Хотя мы исключаем роль положения клетки или ее идентичности в переходе от унификации размера к увеличению вариабельности, мы обнаруживаем, что униформализация размера может, в принципе, наблюдаться в любом клоне вскоре после того, как он инициируется из одной клетки. Одно из неожиданных наблюдений заключалось в том, что гетерогенность роста между клетками внутри клона коррелирует с более быстрым ростом клона.
Зависимость униформизации размера от времени начала клона может быть интересна с точки зрения зависимости от масштаба, потому что униформализация размера наблюдается в меньшем клеточном масштабе, пока клоны не достигнут большего размера, т.е.е. надклеточная шкала — через 24-60 ч, в течение которых начинает проявляться другой ростовой механизм увеличения изменчивости размеров. Это означает, что поведение роста в меньшем масштабе может отличаться от поведения в более крупном масштабе. Чтобы полностью понять этот многомасштабный феномен, можно было бы исследовать рост с постепенным изменением пространственного масштаба и извлечь статистический закон коэффициента роста как функции масштаба. Этот тип анализа роста должен быть ключевой перспективой для понимания роста на надклеточном уровне.
Традиционное значение компенсации в биологии развития растений состоит в том, что баланс между делением клеток и ростом клеток приводит к постоянному размеру органа у мутантов или трансгенных линий в результате изменений размера клеток для компенсации изменений количества клеток в орган (Marshall et al., 2012; Tsukaya, 2003). Здесь мы обнаруживаем аналогичный тип компенсации, то есть униформизацию размера у дикого типа в масштабе клонов.Недавно униформизация размеров наблюдалась внутри сестринских клеток в едином клеточном цикле апикальной меристемы побегов (Willis et al., 2016). Отметим, что в нашем случае униформализация размеров наблюдалась за пределами определенного клеточного цикла с гораздо более длительным временным диапазоном, охватывающим несколько стадий цветения.
Наши результаты долгосрочных временных корреляций (рис. 2) предполагают, что клон все еще сохраняет память о своем первоначальном размере даже после долгого времени.Привлекательная (но все же спекулятивная) гипотеза состоит в том, что контур клона (то есть начальные стенки, включая нижнюю и верхнюю стенки) дает долгосрочные сигналы к росту, поскольку они передаются по наследству. Таким образом, кажется, что клон будет вести себя как отдельная большая клетка и действовать как маленький организм, который помнит свой первоначальный размер.
Наблюдаемая корреляция между ростом клона и неоднородностью роста подразумевает, что сдвиг скорее зависит от восприятия параметров, связанных с ростом.Это поднимает вопрос о точном характере этой реплики. Неоднородность роста порождает механические конфликты (Rebocho et al., 2017), и растительные клетки способны реагировать на такие механические сигналы (Hervieux et al., 2016; Uyttewaal et al., 2012). Следовательно, наш анализ может соответствовать сценарию, в котором способность реагировать на механические сигналы регулируется во времени, делая возможной униформизацию во время развития чашелистника.
Образцы пяти чашелистиков дикого типа были взяты из разных лабораторий (wt-a1, wt-a2: два чашелистика, отображаемые каждые 12 часов в ENS Lyon, Франция; wt-b1, wt-b2, wt-b3: три чашелистика, отображаемые каждые 24 часа. h в Max Planck Cologne, Германия).Мы повторно проанализировали данные, уже использованные в Hervieux et al. (2016), в которых была количественно оценена средняя модель роста чашелистиков.
В сериях a1 и a2 мы использовали линию pUBQ10 :: myrYFP , любезно предоставленную Раймондом Вайтманом (лаборатория Сейнсбери, Кембриджский университет, Великобритания). В этой линии myrYFP соответствует YFP, который на N-конце модифицирован коротким пептидом, который миристоилирован и, вероятно, ацилирован (Yang et al., 2016). Растения выращивали в условиях длинного дня. Стадия определялась, как указано в Smyth et al. (1990). С растения срезали стебли основного соцветия (длиной 1-2 см). Для доступа к молодым бутонам рассекали первые 10-15 цветков. Молодые почки были визуализированы с помощью конфокального лазерного сканирующего микроскопа SP8 (Leica) с использованием дальних 40x (NA 0,8) водно-погружных объективов. Во время покадровой визуализации растения содержали наполовину в среде Мурашиге и Скуга с защитной средой для растений (1 мл / л) и снимали изображения каждые 24 часа в течение до 8 дней.
В сериях b1, b2 и b3 ранние стадии развития цветков (от 3 до 6) были определены на основе сравнения морфологии цветковых бутонов со стадиями, предложенными Smyth et al. (1990). Время развития от стадии 3 до 6 соответствовало времени, опубликованному для аналогичных условий роста (Smyth et al., 1990). Таким образом, более поздние стадии развития тех же цветов были определены путем сравнения времени, начиная со стадии 6 (закрытие бутона), со временем развития, о котором сообщают Смит и его коллеги.
Для исследования роста клеток мы использовали приложение с открытым исходным кодом MGX для визуализации и анализа данных флуоресценции (Barbier de Reuille et al., 2015). Как описано Willis et al. (2016), ключевым преимуществом MGX является способность суммировать данные трехмерной флуоресценции в виде изображения изогнутой поверхности. По данным флуоресценции MGX позволяет нам обнаруживать внешнюю поверхность, указывающую на трехмерную форму органа.После создания поверхностной сетки, которая представляет собой совокупность небольших треугольников вместе с поверхностью органа, мы также можем указать положение клеточных стенок (контуры клеток в основном тексте), используя флуоресценцию мембраны, которая имеет сильную яркость на клеточных стенках. Затем мы можем вычислить удельную площадь поверхности ячейки, чтобы суммировать общую площадь маленьких треугольников внутри ячейки. У нас также есть информация о происхождении, объединяющая набор данных, наблюдаемых в разное время. В этом исследовании мы сначала обнаружили контур клетки на первом временном интервале и отслеживали контур, в котором клетки имеют то же происхождение, что и исходная клетка.Рост клона можно рассчитать как общую площадь клеток клона в момент времени t + Δ t по сравнению с общей площадью клеток клона в момент времени t , то есть A t + Δ т / A т .
Мы рассчитали первый и третий квартили CV, используя метод бутстрапа. Метод начальной загрузки можно кратко охарактеризовать как «случайная выборка с заменой», что позволяет нам измерить точность статистики.Для общего описания предположим, что у нас есть наблюдаемый ценный X (номер выборки N) и его CV (стандартное отклонение X / среднее значение X). Сначала мы создаем «начальный набор данных X B ». То есть мы случайным образом отбираем образцы N раз из набора данных X с заменой и устанавливаем его как X B-1 для первого испытания. Продолжаем таким же образом и создаем наборы данных (X B-1 , X B-2 ,…, X B-1000 ). Используя эти наборы данных, мы можем получить набор «самонастраиваемых CV» (CV B-1 , CV B-2 ,…, CV B-1000 ).Затем мы вычисляем первый и третий квартили распределения бутстрапированной CV.
Используя информацию о темпах роста, мы построим простую феноменологическую модель следующим образом. Основываясь на предыдущей работе (Tauriello et al., 2015), мы аппроксимируем кривую роста площади A t сигмоидальной функциональной формой, т.е. f ( t ) = ( K ). / (1 + exp r ( t c — t ))) + C (рис.S2A). В этой установке скорость роста f ′ ( t ) представляет собой комбинацию линейной и экспоненциальной функций. Из подгоночных параметров (рис. S2B) мы получаем модель как функцию от начального размера, A 0 , как , где γ K = 264, γ C = 2,04 из подгонки и случайной величины ξ K , ξ C равномерно выбираются из диапазона (–10 4 , 10 4 ) и (–10 2 , 10 2 ) соответственно, что соответствует наблюдаемому диапазону шума.Феноменологическая модель показывает линейную зависимость между и, соответственно (рис. S2C), качественно такую же, как и в предыдущем анализе линейной регрессии (рис. 2D, E). Это означает, что линейная зависимость согласуется с функциональной формой сигмовидной кишки с линейной зависимостью параметра K (общий рост клонов) и параметра C (площадь клона в самом начале). Кроме того, мы подчеркиваем, что сама по себе сигмоидальная кривая роста не приводит к линейной зависимости на рис.2D и E. Линейная зависимость является комбинированным эффектом как сигмоидального роста, так и стохастичности на кривых роста, то есть K ( A 0 ) и C ( A 0 ), описанных выше. , среди разных клонов.
Неоднородность роста клеток G h в момент времени t рассчитывается по CV роста отдельной клетки в пределах определенного клона, где — площадь поверхности отдельной клетки в момент времени t .Предположим, что у нас есть M индивидуальной скорости роста клеток, GR t ( C 1 ), в момент времени t внутри определенного клона C 1 , что у нас есть CV, CV 1 ( C 1 ), для конкретного набора данных клона . Аналогичным образом, если мы применили этот процесс к клонам K , мы получим набор данных CV ( CV 1 ( C 1 ), CV 2 ( C 2 ). ),…, CV K ( C K )).Затем термин «неоднородность роста клеток» в основном тексте определяется как усредненное значение набора данных ( CV 1 ( C 1 ), CV 2 ( C ). 2 ),…, CV K ( C K )).
.